THE COASTAL OCEANS OF SOUTH-EASTERN
AFRICA
JOHANN R. E. LUTJEHARMS
"The Coastal Oceans of South-Eastern Africa (15,W)" by Johann R. E. Lutjeharms
reprinted by permission of the publisher from THE GLOBAL COASTAL OCEAN: THE
SEA - IDEAS AND OBSERVATIONS ON THE PROGRESS IN THE STUDY OF THE SEAS,
VOLUME 14, PART B, edited by Allan R. Robinson and Kenneth H. Brink, pp. 783-834,
Cambridge, Mass.: Harvard University Press, Copyright © 2006 by the President and
Fellows of Harvard University.
In The Sea, Volume 14B, editors: A. R. Robinson and K. H. Brink,
Harvard University Press, Cambridge, MA, pp. 783- 834
2006
2
Chapter 20. THE COASTAL OCEANS OF SOUTH-EASTERN
AFRICA
JOHANN R. E. LUTJEHARMS
University of Cape Town
Contents
1. Introduction to the region
2. Mozambique Channel
3. Region east of Madagascar
4. Northern Agulhas Current regime
5. Southern Agulhas Current regime
6. Future research directions
Bibliography
1. Introduction to the region
The coastal ocean off south-eastern Africa is characterised by at least one common,
coherent aspect: it forms part of what may be considered to be the greater Agulhas
Current circulation and all its components are largely dominated by this current
system. In other respects it is very diverse (Schumann, 1998). It extends from the
tropics to a region adjacent to the Subantarctic. The shelf regions are very narrow in
some distinct parts and quite wide in others (Fig. 20-001). Certain parts of the shelf
regions have been studied fairly intensively, while for other regions there is no data or
information to speak of. The current extent of knowledge on this coastal ocean region
is therefore, relying on the existing data base, very inhomogeneous.
Knowledge on the bathymetry and geology of the region has not changed
significantly since a previous review of this kind (Schumann, 1998), so will only be
dealt with here where it affects other aspects of the nature of the coastal oceans. The
equatorward border of the system lies at the northern mouth to the Mozambique
Channel (Fig. 20-001). This is a useful and generic choice and not just one brought
about by geographical pragmatism. The major influence of the Indian monsoon
system on the dominant ocean currents lies entirely to the north of this border (e.g.,
Ridderinkhof and de Ruijter, 2003). Numerical model studies (e.g., Maltrud et al.,
1998) suggest a residual monsoonal effect on currents in the Mozambique Channel,
but direct current observations to date do not support this. By contrast, the poleward
border of the region coincides with the termination of the Agulhas Current at the
Agulhas Current Retroflection south-west of the southern tip of Africa. The eastern
border of the coastal oceans of south-eastern Africa has to include the waters to the
east of the island of Madagascar. As is described below, the coastal waters of the east
coast of Madagascar are not a distinctly separate system (Cooke et al., 2004) but in
many ways form a coherent part of the greater Agulhas Current system. Nevertheless,
for organisational purposes the coastal oceans of this system can be separated into
four distinctive parts: the shelf regions in the Mozambique Channel, the shelf regions
east and south of Madagascar, the shelf inshore of the northern Agulhas Current and,
last,
3
Fig. 20-001 Bathymetry of the South West Indian Ocean in km (after Dingle et
al., 1987a and Simpson, 1974) and the major circulation features. Shelf regions
shallower than 1 km are shaded; hatching indicates upwelling. Some major place
and feature names used in the text are given.
inshore of the southern Agulhas Current. As will be seen, making this geographic
distinction between the southern and northern parts of the Agulhas Current is
important since the nature of the waters and the circulation on their respective shelves
are dissimilar (viz. Fig. 20-001). This comes about as a result of the different
behaviour of the edge of the Agulhas Current in these two regions. For an
understanding of the characteristics of waters on the continental shelves it is therefore
necessary briefly to describe the existing knowledge on the offshore ocean currents of
the greater Agulhas Current system.
The Agulhas Current is supplied with water from essentially two different sources:
the South Equatorial Current and recirculation in a South-West Indian Ocean subgyre
(Stramma and Lutjeharms, 1997; Fig. 20-002). The greater part (40 Sv out of a total
65 Sv in the upper 1 000 m) comes from the subgyre. This configuration of the basin-
wide circulation has been thought to be reflected in the behaviour and natural history
of marine animals (Heydorn et al., 1978), for example the migrations of leatherback
sea turtles (Hughes et al., 1998). The manner in which the South Equatorial Current
acts as a source for the Agulhas Current is not entirely clear. It was previously
thought that this current bifurcates on reaching the east coast of Madagascar - forming
the southern and the northern branches of the East Madagascar Current. The northern
branch of this current and the remainder of the South Equatorial Current would then
pass the northern tip of Madagascar and move onwards to the east coast of the African
4
Fig. 20-002 Circulation patterns of the South West Indian Ocean. (After
Stramma and Lutjeharms, 1997.) The thickness of the lines denotes the volume
flux in the upper 1 500 m. Thin lines indicate lines of historical hydrographic
stations on which the portrayal is based. The Agulhas Current along south-
eastern Africa is largely supported by recirculation in a South West Indian
Ocean subgyre.
continent. Here a similar split would occur with some of the water passing northwards
into the Somali Current and the rest southward into the Mozambique Channel as the
Mozambique Current.
These two western boundary currents - the southern limb of the East Madagascar
Current and the Mozambique Current - would then have a confluence somewhere off
South Africa to form the Agulhas Current. This is the classical portrayal of flow in
this ocean region (Michaelis, 1923) still found in many textbooks and most atlases
and is largely based on analyses of ships' drift (e.g., Sćtre, 1985; Lutjeharms et al.,
2000b). However, modern observations have shown this depiction to be largely
incorrect. The new findings on these currents are of substantial importance for
understanding the flow patterns on the adjacent continental shelves.
It has been demonstrated that no coherent, unbroken western boundary current
exists in the Mozambique Channel. First adumbrated by using the full set of non-
synoptic hydrographic observations (Sćtre and Jorge da Silva, 1984) as well as by
modelling (Biastoch and Krauß, 1999), modern synoptic observations (De Ruijter et
al., 2002) have unequivocally shown that mesoscale eddies are formed at the narrows
of the channel and progress from here southward. No continuous current exists. On
averaging the motions of the drifting eddies (as happens by plotting the means of
many ships' drift observations; e.g., Lutjeharms et al., 2000b) there is the suggestion
of a consistent and continuous boundary current, leading to the classical
misinterpretation.
These Mozambique eddies drift southward along the shelf edge at speeds of
about 5 cm/s and may eventually reach the Agulhas Current (Schouten et al., 2002).
They form the major elements of the circulation in the Mozambique Channel. The
movement in the rest of the channel seems to be sluggish and quite variable, but there
are few observations to support any firm conclusion in this regard, particularly on the
eastern side of the channel. Here there is a dearth of modern hydrographic or current
meter observations (Lutjeharms, 1977). In contrast to classical portrayals, the
Mozambique Channel is therefore now seen as a minor source of water for the
Agulhas Current proper (Fig. 20-002), and only in an intermittent manner. The same
holds for the East Madagascar Current.
5
Instead of being a direct tributary to the Agulhas Current, the southern branch of
the East Madagascar Current has been noted to retroflect south of Madagascar both in
hydrographic data (Lutjeharms et al., 1981a) as well as in satellite remote sensing
(Lutjeharms, 1988). This implies that this current, as in the case of the purported
Mozambique Current, contributes very little to the Agulhas Current, and if so, only
spasmodically. However, the retroflection of the East Madagascar Current has been
shown to be a region that generates both cyclonic as well as anti-cyclonic eddies (De
Ruijter et al., 2003), carrying away some of the shelf waters in the process. Although
these two currents systems East Madagascar Current and Mozambique eddies -
therefore do not form a continuum with the Agulhas Current itself, they do influence
its behaviour indirectly and can therefore be considered to constitute an inherent part
of the greater Agulhas system.
The Agulhas Current proper is fully constituted somewhere near 28° S, along the
east coast of the province of KwaZulu-Natal of South Africa, between Maputo and
Durban (Fig. 20-001). Here the continental shelf is narrow and the course of the
current very stable. This is true for the whole of what might be considered the
northern Agulhas Current (Gründlingh, 1983) extending downstream as far as Port
Elizabeth, at the eastern end of the broad shelf region south of Africa known as the
Agulhas Bank (Fig. 20-001). The coincidence of a narrow shelf and a very stable
juxtapositioned current is not fortuitous; De Ruijter et al. (1999a) have demonstrated
that the continental slope plays an important role in stabilising the trajectory of the
current. The only part of the shelf that does not comply with these stabilising
requirements lies just upstream of Durban, the Natal Bight. At this coastal offset the
shelf is anomalously wide and the continental slope considerably gentler than at other
locations adjacent to the north Agulhas Current. Perturbations on the current path that
occur here will grow, move downstream and have considerable effects on the
subsequent behaviour of the current (Van Leeuwen et al., 2000). This intermittent
meander on the Agulhas Current (Lutjeharms and Roberts, 1988) is called the Natal
Pulse and has a considerable effect on the circulation of the adjoining shelf region.
When the waters in the Agulhas Current reach the Agulhas Bank, the nature of the
current changes dramatically.
From the latitude of Port Elizabeth it is know as the southern Agulhas Current
and, in contrast to the northern Agulhas Current, a range of meanders are formed on
its shoreward side (Lutjeharms et al., 1989a). These meanders cause shear edge
eddies and attendant plumes that move with the current and influence the upper
waters of the shelf in this region. The average trajectory of the current follows the
shelf edge, thus lying farther and farther from the coastline (viz. Fig. 20-001), until
the tip of the Agulhas Bank is passed. In the western lee of the shelf this major current
generates an intense cyclonic eddy (Penven et al., 2001a) that eventually drifts off
into the South Atlantic Ocean. The current itself continues south-westward into the
South Atlantic until it retroflects.
The Agulhas Retroflection is a region exhibiting some of the highest levels of
mesoscale variability in the world ocean (Lutjeharms and van Ballegooyen, 1988;
Garzoli et al., 1996). This is due to the manner in which the retroflection loop
occludes, intermittently forming large Agulhas rings (Lutjeharms and Gordon, 1987)
that then drift off into the South Atlantic (Duncombe Rae, 1991; Duncombe Rae et
al., 1996; Schouten et al., 2000). It has been suggested (Duncombe Rae et al., 1992)
that these rings may interact with the coastal upwelling on South Africa's west coast,
but analyses of the tracks of such rings (e.g., Schouten et al., 2000) indicates that such
6
interaction must be a rare exception. Most rings spend some time in the vicinity of
where they have originally been shed (Boebel et al., 2003), losing a substantial part of
their characteristic heat and salt (Arhan et al., 1999) and having their water masses
exchanged with other rings (Fine et al., 1988) and with Agulhas cyclones (Lutjeharms
et al., 2003b). There is some evidence that Agulhas rings may often be found next to
the western edge of the Agulhas Bank (Lutjeharms and Valentine, 1988) and may
play a role in carrying surface water directly from the Agulhas Current northward past
the western edge of the bank (Lutjeharms and Cooper, 1996).
On moving away from their source region, Agulhas rings tend to move further
away from the African coast (Byrne et al., 1995; Schouten et al., 2000).
Comprehensive reviews of what is currently known about the inter-ocean exchange
that takes place here can be found in Lutjeharms (1996) and in De Ruijter et al.
(1999b). That part of the flow not involved in ring formation, the Agulhas Return
Current (Lutjeharms and Ansorge, 2001), moves water eastward along the Subtropical
Convergence. These deep-sea components of the greater Agulhas system therefore no
longer have a direct influence on the shelf regions. They, as well as the Agulhas
Current proper, may have an indirect effect via their influence on the atmosphere and
on biota.
The Agulhas Current has a marked, visible effect on the overlying atmosphere
through the creation of cumulus clouds (Lutjeharms et al., 1986b), especially along its
southern part and at the Agulhas Retroflection where the heat and moisture loss to the
atmosphere is substantial (Mey et al., 1990). In these regions, under the right
conditions, the thermodynamic considerations adequately account for the formation of
cumulus clouds (Lee-Thorp et al., 1998, 1999). The uptake of moisture in the marine
boundary layer (e.g., Jury and Walker, 1988) can have a marked effect on the
moisture of the adjacent coastal zone and in consequence on the intensities of local
storms and thus on rainfall (Rouault et al., 2002). Increasing rainfall may in turn be
felt in the salinity, water column stratification and colour of the adjacent shelf waters.
Jury et al. (1993) have in fact shown that the distance of the Agulhas Current offshore
has a noticeable effect on the coastal rainfall. Under different climate change
scenarios, the behaviour of the Agulhas Current may change significantly and the
effect on coastal and shelf rainfall may change in concert (Lutjeharms and de Ruijter,
1996).
The Agulhas Current not only has a direct effect on rainfall over the adjacent
coast and shelf, it also has an effect on regional atmospheric circulation patterns
(Reason, 2001) and thus on rainfall over much wider areas. An increase in sea surface
temperature of 2 °C at the Agulhas Retroflection (Crimp et al., 1998) can substantially
affect the atmospheric circulation over the whole southern African subcontinent. An
increase in sea surface temperature over the South Indian Ocean will statistically lead
to an increase in rainfall over regions that form the drainage regions for some of the
main rivers of the South African east coast (Walker, 1990). The effect of increased
runoff on different shelf seas may be very uneven. An investigation of the shelf
waters in the Natal Bight shortly after a major rainfall event (Lutjeharms et al., 2000c)
suggests that it might be relatively insignificant here and restricted to a small area just
offshore of the mouth of the river. Along the Mozambican coast it may, by contrast,
affect the surface salinities of substantial parts of the shelf (Sćtre and de Paula e
Silva, 1979).
To summarise: the south-western Indian Ocean may be considered to be that part
of the South Indian Ocean that is not directly influenced by the monsoonally driven
7
ocean currents. The shelf regions of this particular region are largely dominated by
what may be considered to be the greater Agulhas Current system. Along the east
coast of Madagascar and the east coast of South Africa this consists of well-
developed western boundary currents. Along the east coast of Mozambique it consists
of a series of eddies drifting poleward and along the west coast of Madagascar it may
be a sluggish flow with no distinct pattern or temporal behaviour. Too few
observations are available to characterise the latter unambiguously. This is also true
of the behaviour of shelf waters in many other parts of the Mozambique Channel.
2. Mozambique Channel
A proper understanding of the nature of the waters over the shelves of the
Mozambique Channel is constrained by the large degree of ignorance on the water
movement in the adjacent deep sea. As noted above, historically the flow here has
been visualised as consisting of an intense western boundary current along the east
coast of Mozambique. Although this has now been shown to be incorrect (e.g.,
Ridderinkhof et al., 2001), the average drift patterns at the sea surface (e.g., Sćtre,
1985) nevertheless indicate a strong movement poleward along the eastern shelf of
Mozambique (Fig. 20-004) whereas elsewhere in the channel the average flow is
small and its direction indistinct. The variability of the flow is very high in the
western side of the channel, but low in the eastern side. This variability is evident in
analyses of ships' drift (Lutjeharms et al., 2000b), altimetric observations (e.g.,
Lutjeharms et al., 2000d) and in modelling of the region (e.g., Biastoch and Krauß,
1999). These results both strong currents at the shelf edge and high variability of
course all agree with the concept of a train of eddies moving poleward through the
channel. Knowledge on the deep sea circulation that may affect the shelves is even
more lacking on the eastern side of the channel.
Ships' drift observations, altimetry and even the few hydrographic observations
give no unambiguous indication of the movement of water here. In general speeds are
low and to the north in the ships' drift observations (Sćtre, 1985; Lutjeharms et al.,
2000b) whereas modelling results (e.g., Biastoch et al., 1999) indicate an average
flow to the south. Interpretations from the very few hydrographic data (e.g., Menaché,
1963; Sćtre and Jorge da Silva, 1984; Donguy and Piton, 1991) have included a
northward flow as well as a southward flow. The only known direct observations
indicated a weak northward (Martin et al., 1965) and a weak southward flow (Piton
and Poulain, 1974). Satellite observations have indicated the presence of cyclonic
eddies off the shelf edge at the south-western coast of Madagascar that draw coastal
water, rich in chlorophyll-a, seaward (Quartly and Srokosz, 2004). What does stand
out in all appropriate data is that the intensity of current variability on the eastern side
of the channel is considerably lower than on the western side.
2.1 General water mass characteristics in the channel
The temperature-salinity characteristics of the waters in the Mozambique
Channel are shown in Fig. 20-003. This figure is based on relatively old hydrographic
data, but homogeneously covers a greater part of the channel than more modern data.
It demonstrates that the salinities of waters in the upper layers lie in a band between
8
35.00 and 35.40, with a few outliers, mostly as fresher water. These particular outliers
may be due to river runoff. At 18° C there are two different water masses, identifiable
Fig. 20-003 The temperature-salinity and temperature-oxygen characteristics
of the water masses in the Mozambique Channel. (After Lutjeharms, 1991.)
Sigma-t isolines have been added on the T-S scattergram. Water types that are
evident are Equatorial Indian Ocean Water (Eq IOW), Subtropical South Indian
Ocean Water (Subt SIOW), Central Water, Antarctic Intermediate Water
(Antarc IW), North Indian Deep Water (NIDW) and North Atlantic Deep Water
(NADW).
by distinct salinities. They are Equatorial and Subtropical Indian Ocean Water
respectively and are found in different parts of the channel on different occasions. No
durable delimitation for the distribution of either is to be found (Sćtre and Jorge da
Silva, 1984), although Equatorial Indian Ocean Water (also called Tropical Surface
Water) is found largely in the northern part of the channel. Since waters to a depth of
900 m are know to upwell onto the Mozambican shelf (e.g., Lutjeharms and Jorge da
Silva, 1988) one can expect to find water types down to the less saline water of the
Central Water (viz. Fig. 20-003) in some coastal regions, but most of the waters on
shallow shelves would come from waters in more superficial layers.
9
Fig. 20-004 Bathymetry of the Mozambique Channel and the continental shelf
off Madagascar in km (after Simpson, 1974) with the major circulatory features.
Shaded areas are shallower than 1 km; hatched areas denote upwelling. Names
of places and features mentioned in the text are given.
2.2 Effects of shelf morphology
The general bottom morphology of the shelves in the Mozambique Channel is shown
in Fig. 20-004. The shelf is narrow on the western side of the narrows of the channel
at about 16° S, but wide on the eastern side. They are fairly narrow on both sides of
the channel at its southern mouth. The rest of the Mozambican shelf is wide, while
that off Madagascar is narrower. Just south of the mouth of the channel there is an
extensive offset in the coastline just off the Mozambican capital of Maputo, the
Delagoa Bight. There is also an offset south of Angoche (Fig. 20-004). The shelves
around the numerous islands, particularly the Comores in the northern mount of the
channel, are narrow. These shapes of the shelf edge have a decided effect on the
coastal water movement.
One of the consequences of the changes in direction of the coastline is in the
formation of coastal lee eddies. An example of such a feature can be seen off the town
of Angoche (Fig. 20-005). Here the flow along the greater part of the shelf edge - and
probably on the shelf as well - is strongly poleward. At the time it was assumed that
this formed part of a continuous Mozambique Current (Nehring, 1984); currently the
10
consensus is that this most probably was part of the edge of an anti-cyclonic eddy
drifting southward (e.g., De Ruijter et al., 2002). Notwithstanding the ignorance on its
Fig. 20-005 Lee eddy at Angoche along the Mozambican coast (viz. Fig. 20-
004). The dynamic topography of the sea surface relative to the 600 dbar level is
given in dynamic centimetre, based on a cruise undertaken in 1980. Dots
represent station positions. The continental margin shallower than 1 km is
shaded. (After Nehring et al., 1987.)
source, the important thing to note is the fact that there was a strong current poleward
on this occasion, even though it might have been intermittent. This current overshot
the offset in the coast at about 16° S, forming a distinct lee eddy to the south (Fig. 20-
005). This eddy had a diameter of about 100 km and deeper water was upwelled in its
core (Schemainda and Hagen, 1983). This is shown by an enhanced nutrient content
of more than 12 mol/ nitrate-nitrogen at 75 m depth compared to 2-4 mol/ in the
offshore current at the same depth. A resultant peak in chlorophyll-a concentration
was also observed in this lee eddy (Nehring et al., 1987). Although no hydrographic
stations were carried out on the shelf itself, the implication clearly is that the motion
on the adjacent shelf would be equatorward.
The shelf configuration at this presumed lee-eddy is similar to that of the St Lucia
and the Port Alfred upwelling cells (Lutjeharms et al., 1989b; Lutjeharms et al.,
2000a; Figs 20-001 and 20-016). At all three of these locations a strong current along
the shelf edge moves past an offset in the coastline, from a narrow shelf and then past
a wider shelf. According to the theory put forward by Gill and Schumann (1979), this
should lead to upwelling inshore of the strong current. In the former two cases it does,
therefore perhaps also off Angoche at about 16° S in the Mozambique Channel. In his
analysis of seasonal sea surface temperatures and the depths of isotherms along this
coastline, Jorge da Silva (1984a) has shown the prevalence of upwelling at this spot,
but only for certain periods of the year. His database for such seasonal analysis was
not very large, so that this result can only be considered tentative. If the hypothesis is
correct and this upwelling is driven by poleward currents as part of passing eddies, it
would be sporadic and not a continuous feature. Nevertheless, this spot represents the
11
highest observed chlorophyll-a values along this coastline (Nehring et al., 1987) and
may therefore have a decided influence on the ecosystem of this whole shelf region.
If the offshore eddies that form at the narrows (De Ruijter et al., 2002;
Ridderinkhof and de Ruijter, 2003; Fig. 20-004) consistently move along the shelf
edge it is likely that the waters on the adjacent shelf would experience alternating
poleward and equatorward setting currents. This situation is also found on the shelf
adjacent to the northern Agulhas Current where the shelf waters move largely in
harmony with the current, but on the intermittent passing of a Natal Pulse, reverse and
set strongly against the direction of the Agulhas Current (Lutjeharms and Connell,
1989). Quartly and Srokosz (2004) have used satellite observations of ocean colour to
demonstrate that passing Mozambique eddies extract water from the neighbouring
Mozambican shelf and inject it into the mid-channel region. In this way the exchange
of waters between the shelf and the deeper part of the channel will consist of rather
frequent episodes, driven from afar. They (Quartly and Srokosz, 2004) have also
shown that the passage of eddies past the Delagoa Bight appears to affect the
circulation there.
Fig. 20-006 The Delagoa Bight eddy off the city of Maputo in southern
Mozambique (viz. Fig. 20-04). Isotherms at 200 m depth show the cold water
upwelled in the centre of the eddy, based on a cruise undertaken in 1982. (After
Lutjeharms and Jorge da Silva, 1988.) The broken line shows the intersection of
the 200 m isobath with the shelf edge; the shelf itself is shaded.
The Delagoa Bight is a large offset in the coastline at the latitude of Maputo (see
Fig. 20-004). The flow past here, either as the start of the Agulhas Current or as the
landward end of passing, anti-cyclonic Mozambique eddies, is poleward. This passing
water generates a cyclonic flow in the bight (Lutjeharms and Jorge da Silva, 1988)
and the resultant Delagoa Bight eddy dominates the flow at the shelf here throughout
most of the year (Fig. 20-006). Over a period of 23 years it has been observed 11
times in the hydrographic results of research cruises. It has been hypothesised that this
12
recurrent eddy has largely determined the distribution of the sediment base of the
Delagoa Bight (Martin, 1981). There is evidence (Gründlingh, 1992a) that these
eddies may on occasion escape from the bight and drift into the deep ocean. On this
part of the Mozambican shelf the water movement will most probably be totally
dominated by this lee-eddy. If the same effects are found here as in lee-eddies in the
other offsets in this coastline (e.g., Nehring et al., 1987), it can be assumed that there
is considerable vertical movement of water in the lee eddy (Schemainda and Hagen,
1983). Water masses in this eddy have temperature-salinity characteristics implying
substantial upwelling in the core of the eddy from depths of at least 900 m. Hence
there should be nutrient enrichment of the surface layers and thus increases in the
chlorophyll-a content. To date this has not been observed, except intermittently at the
north-eastern point of the Delagoa Bight (Quartly and Srokosz, 2004). This sporadic
increase in primary productivity may be the result of current-induced upwelling as
predicted by Gill and Schumann (1979). If driven by passing eddies, such intermittent
upwelling would be expected.
The shelf morphology of the eastern part of the Mozambique Channel is different
to that of the western part (Fig. 20-004). For the most part the shelf is narrow except
at the narrowest part of the channel at about 17° S where the eastern shelf is
widest. As mentioned before, the flow along this eastern shelf edge is quiescent
compared to the western side of the channel (Sćtre, 1985; Lutjeharms et al., 2000b)
with low speeds and low eddy kinetic energy. The average current direction seems to
be equatorward. Nevertheless, there are tentative indications from the distribution of
chlorophyll-a (Quartly and Srokosz, 2004), as observed from satellite, that cyclonic
eddies form along the southern tip of Madagascar and drift into the channel. They
seem to draw water off the shelf and inject it into the waters of the central channel.
In summary, the shelf edges of the western part of the Mozambique Channel
seem to be influenced mainly by passing Mozambique eddies that have their origin in
the narrows of the channel and by secondary effects due to these eddies, namely
upwelling at coastal offsets, the generation of lee eddies at shelf offsets and the
extraction of shelf waters to mid-channel. It is possible that parts of the eastern shelf
of the Mozambique Channel are also influenced by eddies, but the data at hand are
inadequate to show this unambiguously. It is evident that such eddies would be
considerably more infrequent than and not as intense as those that affect the western
shelf of the channel. Besides the direct influence of the currents at the shelf edge, the
waters over the shallower parts of the shelf may be substantially influenced by the
reigning winds.
2.3 Effect of winds and tides
The meteorological conditions for the Mozambique Channel have been summarised
by van Heerden and Taljaard (1998). In austral summer the mean wind direction is
uniformly from a south-easterly direction and weak. In winter the average wind
direction differs for the southern and the northern part of the channel. The southern
part experiences south-easterly winds; the northern part north-westerlies. The
northern part thus may be considered to form part of the monsoonal wind system of
the Indian Ocean up to 15° S (Sćtre and Jorge da Silva, 1982) whereas the southern
part does not. The border between these two systems is the Inter Tropical
Convergence Zone. Donguy and Piton (1991) have attempted to relate the currents in
the channel to the monsoonal winds, but with only a few cruises and short sea level
13
records at their disposal a conclusion of monsoonal seasonality in the currents is
probably premature. As mentioned above, the large scale current systems show no
evidence of monsoonal influence.
The winds over the shelf regions agree in part with the current direction
hypothesised by Sćtre and Jorge da Silva (1984). Except for the most southern part of
the western shelf the Delagoa Bight the winds along this coastline all have a
strong equatorward component year round. Sćtre and Jorge da Silva (1984) have
therefore concluded that the shelf currents here also are in that direction. These
inferred current directions are in contrast to ships' drift observations (Sćtre, 1985;
Lutjeharms et al., 2000b) that show consistent movement poleward over the western
shelf region. The winds over the eastern shelf are also largely northward, but vary
with season.
During summer the shelf waters of the Mozambique Channel may also be affected
by passing tropical cyclones that, as a rule, move poleward through the channel (Van
Heerden and Taljaard, 1998). Some, however, make landfall somewhere along the
coast of Mozambique. The effect of passing cyclones on the water movement over the
shelves is dramatic. In situ current observations have shown (H. Ridderinkhof,
personal communication) a reversal of current direction to a depth of at least 200 m at
the passing of a severe cyclone.
Tides are an important component of water motion in the Mozambique Channel,
especially when compared to other parts of the South-West Indian Ocean where tidal
ranges are small. There is a gradual increase in tidal range from less than 2 m to the
north and to the south of the channel to more than 5 m in its central part. This is the
product of a double standing wave system, driven from either end, which develops in
the channel (Pugh, 1987). Low lying coastal regions and estuaries, particularly on the
Mozambican side, contain extensive areas of salt marshes and mangrove swamps as a
result of the tidal motion (G. B. Brundrit, personal communication). Over the wide,
shallow parts of the Sofala Bank (viz. Fig. 20-007) strong tidal currents lead to the
continuous movement of sand banks and other mobile sedimentary seabed features.
In summary, tidal currents are important in the shallow parts of the shelves, but the
usually weak winds have a limited affect on the main water movement over the
shelves of the Mozambique Channel, with the exception of passing cyclones whose
influence might be short lived.
Apart from the wind regimes, in a region of high rainfall (Van Heerden and
Taljaard, 1998) the influence of river runoff on the shelf waters may be important.
2.4 Effect of river runoff
The runoff from the Mozambican land mass varies seasonally, but there also are
considerable interannual differences (Jorge da Silva, 1984c). The total runoff from the
Zambezi River (viz. Fig. 20-004, 20-007), for example, was 168.9 km3 for 1977-78;
whereas for 1982-83 it was a mere 50.3 km3. Intermittent tropical cyclones bring
event-scale rainfall episodes that may totally dominate the annual rainfall distribution.
This naturally also holds for the Madagascar land mass, but runoff records for that
country are difficult to obtain. One of the few coastal regions for which accurate
observations of the effect of the river runoff on the shelf waters have been made on a
number of occasions (Jorge da Silva, 1984c; 1984b) is along the Sofala Bank. It lies
between 16° and 21° S on the western shelf of the channel (Fig. 20-004).
14
The salinity of surface waters close to the Zambezi River mouth on the Sofala
Bank may drop as low as 20.00 (Sćtre and de Paula e Silva, 1979) at a time when
water at the shelf edge may be 35.4 (viz. Fig. 20-007). This river water may be
severely discoloured, giving a Secchi disc depth of less than 2 m (Jorge da Silva,
1984b). The
Fig. 20-007 Surface salinities on the Sofala Bank - the wide, shallow shelf off
central Mozambique - based on a cruise of 1982. (After Jorge da Silva, 1984c.)
For general location see Fig. 20-004. Dots show station positions. Locations of
station sections in the bottom panels are shown by roman numerals in the upper
panel. The shelf shallower than 50 m is shaded. Fresher water (< 35.0) extended
to a depth of 15 m directly off the Zambezi River mouth; to 30 m off the Luala
15
River mouth. Note the high salinity values south of the city of Beira, due to
marsh runoff.
plume of muddy water can be quite extensive (Sidorn et al., 2001) and has been
thought to play a major role in the natural history of local biota, such as shrimp. The
fresher water may extend over the shelf to a distance of 50 km offshore. It may be
confined to the top 15 m of the water column, or it may extend to the full depth of the
shelf (Fig. 20-007), presumably dependent on the density differences between the
river and shelf waters as well as the concurrent wind action. Plumes of fresh river
water have been observed to extend both equatorward as well as poleward from most
rivers here. No clear pattern of movement is therefore immediately evident. Sćtre and
de Paula e Silva (1979) have shown that the greater part of the Sofala Bank is affected
by fresher surface water.
The chlorophyll-a distribution as well as the primary productivity on the Sofala
Bank seems to be largely controlled by the effluent from rivers. These carry loads of
nutrients (Jorge da Silva, 1984c) that, as mentioned above, may penetrate half-way
across the shelf. The chlorophyll-a distributions exhibit very analogous patterns. That
these distributions occur with a high frequency can be seen by the organic matter
content of the surface sediments on this part of the shelf. It is highest directly off the
Zambezi River mouth and in a narrow strip adjacent to the coast to either side of this
river mouth. Some small pelagic fish seem to concentrate in these waters (Jorge da
Silva, 1984c). Otherwise pelagic fish on the Sofala Bank seem chiefly to frequent
strips parallel to the coast, most extending along the whole coastline (Brinca et al.,
1981).
One may quite reasonably expect the observed effects of fresh water runoff on the
Sofala Bank to hold for the rest of the shelf regions of the Mozambique Channel as
well, where there are fewer measurements of this kind. The salinity of shelf waters
along most of Mozambique varies seasonally with the river outflow, with the lowest
salinities found in February (Sćtre and de Paula e Silva, 1979). Apart from the
expected seasonality, results for the Sofala Bank show great inter-annual variability
and one could assume that this holds for all the other Mozambican shelf regions as
well. The freshening of the shelf waters off Mozambique by river run-off seems
ubiquitous.
There is at least one clear exception. At the southern extremity of the Sofala Bank,
between 20° to 21° 30' S (viz. Fig. 20-007), there is a large expanse of coastline that is
subject to inundation by seawater. The subsequent runoff from this region can be
extremely salty. Salinity values in excess of 36 are not uncommon (e.g., Jorge da
Silva, 1984c). During September of 1982 surface values of 37.2 were observed near
the coast (Brinca et al., 1983). The influence of this saline water has been observed a
distance of 100 km offshore and throughout the shallow water column of 50 m.
2.5 Characteristics of the western shelf
To recapitulate, the water mass characteristics of the shelf waters on the western shelf
of the Mozambique Channel have been presented by Jorge da Silva et al. (1981),
Jorge da Silva (1984a), Lutjeharms and Jorge da Silva (1988) and Sćtre and Jorge da
Silva (1982). Except for regions and times where river run-off plays a major role, the
water masses over the shelf are identical to those offshore (viz. Fig. 20-003) at the
same depths. This means that the subsurface salinity maximum of Subtropical Surface
16
Water (at depths of 150 300 m) is much more pronounced in the southern part of the
channel than farther north. In the Delagoa Bight eddy, at the southernmost extremity
of the shelf (Lutjeharms and Jorge da Silva, 1988), there is hardly any evidence of
Tropical Surface Water left. The exchange of water masses between the shelf and the
deep ocean seems to vary considerably. The passage of Mozambique eddies may play
a key role here. At parts of the western shelf where the shelf is very narrow (viz. Fig.
20-004) and the current at the shelf edge strong, such as at the narrows of the channel
(Angoche; Sćtre, 1985) and just north of the Delagoa Bight (Inhambane; viz. Fig. 20-
004; Lutjeharms et al., 2000b), one may expect that the shelf edge currents may have
a more decided effect, whereas along the Sofala Bank where the shelf is widest (Fig.
20-004), this effect would be substantially less.
Sćtre and Jorge da Silva (1982; 1984) have carried out the most detailed analyses
of water masses on this shelf to date. They have claimed that the circulation patterns
of the shelf waters can be visualised by the temperature distribution at 150 m depth.
This leads to a different pattern for the hydrographic results of each research cruise
for the region. A set of cyclonic eddies of various shapes and sizes are evident. Based
on these data one may therefore safely assume that the waters and the circulation on
this shelf region are very variable. What effect does this have on the ecosystem of this
shelf region?
Surveys of organisms and in particular of fish resources have been made on the
western shelf region of the Mozambique Channel (Nehring, 1984; Nehring et al.,
1987; Sćtre and de Paula e Silva, 1979; Jorge da Silva, 1984a), but particularly on the
Sofala Bank (Brinca et al., 1981; Jorge da Silva, 1984c). The values of column
chlorophyll-a concentration are relatively low over most of the outer parts of the
shelf, somewhat higher at the shelf edge (Sćtre and de Paula e Silva, 1979). Over
inner parts it can rise to 98 mg/m2 (Nehring et al., 1987). The exception, mentioned
above, are the observations in the upwelling cell off Angoche where values of 600
mg/m2 have been observed. The distribution of zooplankton biomass in the upper 30
m of the water column shows a similar general distribution with low values of 20-40
mg/m3 over most outer parts of the shelf with higher values, up to 160 mg/m3 at inner
stations. The exception is for the region directly poleward of the Angoche upwelling
cell where observations of 320 mg/m3 have been made. No evidence for such major
increases in either phytoplankton or zooplankton have to date been found in the
Delagoa Bight eddy. Even though Nehring et al. (1987) have shown that the primary
productivity at inshore stations on the shelf was double that of stations farther
offshore, there were considerable differences between stations close to each other on
the shelf. Large degrees of spatial and temporal variability can therefore be assumed.
An analysis of the seasonal distribution of the depth of the 20° and the 23° C
isotherms has shown that large parts of the outer shelf would in principle be suitable
for yellowfin tuna fisheries, whereas the skipjack tuna are most likely to be found off
Angoche and Maputo, i.e. at the sites of upwelling induced by lee eddies (Fig. 20-
008). The distribution of fish; demersal, small pelagic, larger pelagic, mesopelagic as
well as that of crustaceans has been summarised by Sćtre and de Paula e Silva
(1979). In most cases where more than one survey cruise was carried out there was a
considerable difference between the cruise observations and this was usually
considered to be due to seasonality in the distribution of organisms. It may have been
due to irregular temporal changes of shorter duration.
In short, the circulation on the Mozambican shelf is very variable in both space and
time and may be influenced by offshore currents only where the shelf is narrow. Run-
17
off from land plays a key role, but exhibits both seasonal and inter-annual variations.
The biogeography exhibits the same variability. Although the interpretation of the
circulation patterns as well as the biogeography of this western shelf of the
Mozambique Channel is constrained by limited data, this is even more the case for the
eastern shelf.
Fig. 20-008 Tuna vulnerability to catching by surface gear on the Mozambican
shelf, superimposed on average sea surface temperatures during the period
January to March. (After Jorge da Silva, 1984a.) The 50 m isobath is shown.
Note the increased concentration in the lee eddies off Angoche and in the
Delagoa Bight off Maputo.
2.6 Characteristics of the eastern shelf
18
The eastern shelf of the Mozambique Channel can using information currently
available - be divided into three specific regions: the very south where the shelf is
more or less zonal, the very north, where the shelf forms part of the Comoro Basin
(viz. Fig. 20-004) and, third, the meridional shelf in between.
The Comoro Basin lies directly south of the South Equatorial Current (Piton and
Poulain, 1974). Between it and the narrows of the channel an anti-cyclonic gyre is
formed that seems relatively stable (Donguy and Piton, 1991). The surface currents of
which the gyre consists are not strong (Sćtre, 1985) except near the African coast
where mean speeds in excess of 0.5 m/s are to be found (Lutjeharms et al., 2000b).
The variability on the African side is high. The currents over the eastern shelf in the
Comoro Basin are weak but in general in an equatorward direction. This rather
inadequate portrayal is nonetheless supported by direct measurements (Piton and
Poulain, 1974; Martin et al., 1965) as well as by a variety of models (e.g., Biastoch
and Krauß, 1999; Asplin et al., 2004; Sćtre, 1985).
The water types on the shelf are dominated here by Tropical Surface Water with a
salinity range from 34.3 to 35.2 (Donguy and Piton, 1969). There is no evidence to
date that Subtropical Surface Water with greater salinity is found on the shelf,
although in principle it is entirely possible since this water mass is found at about 200
m depth in the region. Surface temperatures exceed 26° C throughout the year, except
in the months of August and September. Highest temperatures (> 29°) are found in
April; lowest (25° to 26° C) in August. Surface salinities are lowest (34.4) in March;
highest in the period September to November (>35.10). Recent observations on this
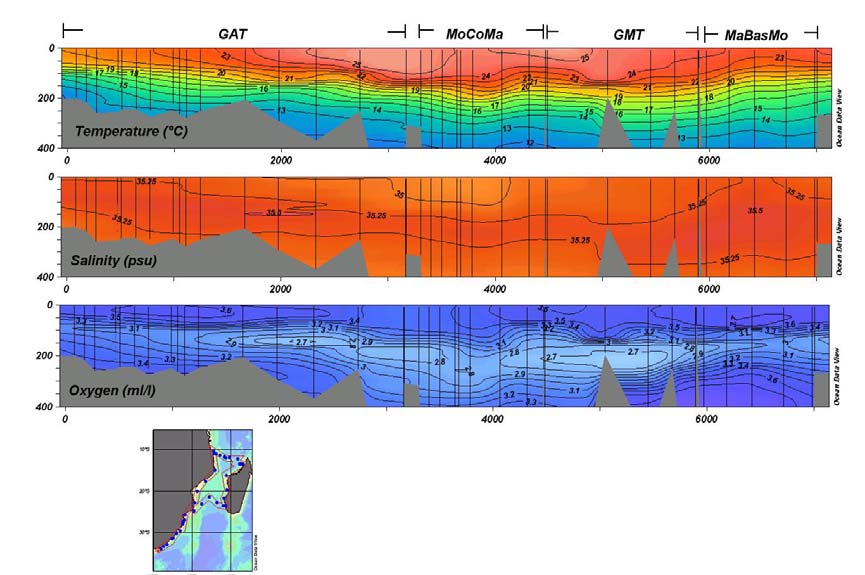
Fig. 20-009 Hydrographic characteristics of shelf waters along the east coast
of South Africa and the continental shelves in the Mozambique Channel. (With
special permission of M. Roberts.) GAT is the section from Port Elizabeth in the
south along the east coast of South Africa and of Mozambique; MoCoMa
represents the zonal section from the coast of Mozambique to the west coast of
Madagascar via the Comores; GMT a poleward section along the west coast of
19
Madagascar and MaBasMo a zigzag section from the southern point of
Madagascar to the island of Bassas da India and on to Maputo on the east coast
of Mozambique.
shelf region (Roberts, personal communication) shows that the water column consists
of a warm mixed layer to a depth of about 80 m with the seasonal thermocline
extending to 250 m. Salinities in the centre of the Comoro Gyre are slightly elevated
above those found on the adjacent shelves. An oxygen minimum lies at 200 300 m
depth and is less strongly developed in the centre of the gyre. The hydrography of the
middle part of the western shelf of Madagascar is not much different.
Here, according to the only observations to be found (Roberts, personal
communication) the warm mixed layer extends to 150 m, with a uniform salinity
between 35 and 35.25 (Fig. 20-009). Particularly noteworthy is the oxygen minimum
layer found between 150 and 250 m that is strongly developed here, more so than
anywhere else on the shelves of the region. How representative these observations are
remains unknown. The little that is known about the currents along the shelf edge
(e.g., Sćtre, 1985; Lutjeharms et al., 2000b) here, suggest that they are very weak and
will most probably have very little influence on the movement of the waters on the
shelf itself. As mentioned before, satellite imagery, particularly of ocean colour
(Quartly and Srokosz, 2004), shows that cyclonic eddies along this coastline may on
occasion draw off substantial amounts of shelf water and with it chlorophyll-a into
the deeper parts of the channel. The origin of these eddies, between 200 to 300 km in
diameter, is uncertain. De Ruijter et al. (2003) have shown that cyclones can be
formed at the termination of the southern East Madagascar Current and these could
Fig. 20-010 The temperature/salinity (left panel) and the temperature/dissolved
oxygen characteristics for the shelf waters south of Madagascar. (After
20
Anonymous, 1983.) The line of stations on which this was based was taken to the
south-west of Andriamanao (viz. Fig. 20-04).
conceivably drift into the Mozambique Channel along the shelf edge. Although a
number of hydrographic interpretations (e.g., Sćtre and Jorge da Silva, 1984; Donguy
and Piton, 1991) imply a southward setting current along this shelf, direct
measurements (Martin et al., 1965) and ships' drift observations (Lutjeharms et al.,
2000b) indicate otherwise. Cyclonic eddies could therefore be advected equatorward
along this coast.
The third and last component of the eastern shelf of Madagascar is the southern
part. The movement of the shelf waters here may be more dynamic, since it could
conceivably be affected by the southern limb of the East Madagascar Current (viz.
Fig. 20-004). This would most likely be true largely for the eastern part of this shelf
region (Lutjeharms et al., 2000b). Hydrographic observations on the shelf show the
normal temperature/salinity characteristics to be expected (Fig. 20-010). Surface
temperatures (in June) were 24 °C and surface salinities 35.3. (The higher
temperatures shown in the scatter diagram of Fig. 20-010 represent temperatures
farther offshore.) Both Tropical and Subtropical Surface Waters are evident in the
temperature/salinity values. The Tropical Surface Water extended from the surface to
about 100 m depth. The Subtropical Surface Water below that extended to about 250
m. A well-developed subsurface oxygen minimum was found at a depth of 80 to 200
m.
The surface salinities and temperatures all suggest an upwelling regime on this
southern shelf. During the first extensive cruise over this shelf (June 1983;
Anonymous, 1983) the temperatures at the coast were 2 °C lower than further
offshore. The salinities were up to 35.6, indicating upwelled Subtropical Surface
Water (see Fig. 20-010). Ocean colour also shows signs of enhanced chlorophyll-a
values along this coastal segment (Quartly and Srokosz, 2004) and that these may be
drawn off the shelf in plumes. The presumed upwelling is concentrated in an
upwelling cell on the south-eastern corner of Madagascar (Lutjeharms and Machu,
2000; DiMarco et al., 2000) where enhanced concentrations of chlorophyll-a and
lower temperatures are found most frequently. The question remains if these remotely
sensed suggestions of upwelling may not be due to runoff from land.
Hydrographic evidence specifically collected to ascertain the origin of these
elevated values of chlorophyll-a (Machu et al., 2002) has recently shown
unequivocally that there is indeed upwelling at this location. The upwelling does not
seem to be strictly related to the wind patterns and it has therefore been hypothesised
(Lutjeharms and Machu, 2000) that the driving force for the upwelling is the
juxtapositioned East Madagascar Current. This would be comparable to the upwelling
at the eastern extremity of the Agulhas Bank (Lutjeharms et al., 2000a; viz. Fig. 20-
016) and at the northern extremity of the Natal Bight (Lutjeharms et al., 1989b; viz.
Fig. 20-011). However, there is some evidence (Quartly and Srokosz, personal
communication) that the chlorophyll-a concentration at this location has a distinct
seasonal pattern with the highest concentrations found in the austral winter months of
July and August, the lowest in December. Major winds at this location are from the
east in winter, from the north-east in summer (Sćtre, 1985) suggesting winds more
favourable for upwelling along the full south coast of Madagascar in winter, at the
south-eastern corner in summer. The importance of the wind compared to the current
in driving this upwelling therefore remains unresolved.
21
The biological implications of this upwelling are intriguing, but to date not
properly quantified. Apart from the remotely sensed chlorophyll-a, surveys of fish
stocks (Anonymous, 1983) have shown slightly higher concentrations of demersal
fish on the southern shelf than off the adjacent, eastern shelf. Mackerel numbers were
higher on the southern shelf, but scad lower. In general fish were found with such a
very scattered distribution on this shelf that no firm conclusions can be reached on
their biogeography.
In summary, the waters over the shelves of western Madagascar seem fairly
unusual, based on the current - very limited - data. Water masses are those found
offshore, except for an intensification of the subsurface oxygen minimum, and the
currents are most probably weak and variable. The wider, southern shelf exhibits
characteristics of upwelling, but this does not seem to have a marked effect on higher
trophic levels.
An outline of the characteristics of the shelf waters of the Mozambique Channel as
a whole is, as can be seen from the above descriptions, largely a function of the
amount of available data and their quality. The broad Mozambican shelf is
characterised by substantial terrestrial influence from runoff both from rivers and salt
marshes. Lee eddies at Angoche and in the Delagoa Bight may play an important, but
local role. The effect of Mozambique eddies intermittently passing by the shelf edge
is not known. Water masses over the northern shelves are predominantly Tropical
Surface Water, those to the south Subtropical Surface Water. This seems true for the
western as well as the eastern shelves of the Mozambique Channel. About the latter
very little is known, except that it is relatively broad.
The shelf to the east of Madagascar is by contrast much narrower and the offshore
circulation totally different.
3. Region east of Madagascar
Not only is the shelf narrow here, but the continental slope is precipitous (viz. Fig. 20-
004). Along the shelf flows a small, but intense western boundary current, the East
Madagascar Current (e.g., Lutjeharms et al., 1981a). Swallow et al. (1988) have
observed a speed in the southern limb of this current of 0.66 m/s, at a latitude of 23
°S, about 50 km from the coast, with a standard deviation of only 12 cm/s. The speeds
in the northern branch of this current (Schott et al., 1988) are not much different. The
separation point between the northern part, flowing equatorward, and the southern
part, flowing poleward, is estimated to lie between 17 and 18 °S (e.g., Lutjeharms et
al., 2000b; viz. Fig. 20-004). This may vary with season as the wind patterns shift
northward along this coastline in the austral winter (Van Heerden and Taljaard, 1998).
No direct observations have been made to date, but one may assume that the waters
over the shelf move in concert with the strong offshore currents.
As can be expected, the temperature/salinity relationships of the waters on this part
of the shelf are indistinguishable (Anonymous, 1983) from those found on the shelf
south of Madagascar (Fig. 20-010).
Little is known about the biological productivity of the region. Fish distributions
are scattered with a decrease in pelagic as well as demersal fish as one moves
equatorward. Catches of sharks and rays increase on going northward along this coast
(Anonymous, 1983).
22
In summary, all that is known with a certain degree of certainty about the shelf
seas east and south of Madagascar is the presence of the East Madagascar Current at
the shelf edge and the likelihood of persistent upwelling south-east of the island. The
interaction between these or the influence of the current on the shelf circulation
remains unknown due to an extreme paucity of observations.
Compared to the lack of data and knowledge about this particular part of the shelf
seas of the South-West Indian Ocean, the region inshore of the northern Agulhas
Current is very much better studied and understood.
4. Northern Agulhas regime
The northern Agulhas Current is defined as that part of the current extending from the
southern mouth of the Mozambique Channel downstream to the eastern edge of the
Agulhas Bank (viz. Fig. 20-001). This component of the current flows past a shelf that
may be considered to consist of two categories. For the greater part the shelf is narrow
and the continental slope has a steep gradient. The only exception is a part of the shelf
along the province of KwaZulu-Natal known as the Natal Bight. Here the shelf is
considerably wider and the slope much broader and with a gentler gradient (Fig. 20-
011). As will be seen below, this shelf morphology has some remarkable effects on
the offshore currents.
23
Fig. 20-011 The bathymetry of the continental shelf along the northern Agulhas
Current. (After Simpson, 1974.) The continental shelf area is shaded. The 200 m
isobath is shown as a broken line. Hatched areas denote upwelling. Place names
and circulatory features are mentioned in the text.
The core of the northern Agulhas Current follows the shelf edge very closely
almost all of the time (Tripp, 1967) meandering less than 15 km to either side
(Gründlingh, 1983). For a western boundary current this is quite unusual, but it has
important consequences for the circulation on the adjacent shelf. As can be expected,
in its very surface layers the behaviour of the current is not as stable (Pearce, 1977a)
with changes in speed occurring from day to day and the penetration of surface water
of the current onto the shelf taking place at irregular intervals. In some cases short-
term current reversals at the edge of the current have been observed (Gründlingh,
1974; Schumann, 1981; Pearce et al., 1978), possibly due to shear edge eddies or to
the effect of the wind. Surface speeds of the inshore edge of the current may exceed
1.5 m/s, salinities may lie between 35.00 and 35.50 and temperatures in summer may
exceed 28. In winter these sea surface temperatures drop to less than 21 °C (Pearce,
1978). Shallow water near the shelf edge is usually Tropical Surface Water. The
characteristic salinity maximum of Subtropical Surface Water is found at a depth of
150 to 250 m at least 60 km offshore (Pearce, 1977a) although this distance may vary
on a near-daily basis. One would therefore expect the waters over the adjacent shelves
to consist largely of modified Tropical Surface Water, but as it turns out, this is not
the case.
This established current disposition is not entirely stable, as mentioned above.
During about 15% of the time and at irregular intervals the current moves offshore
in a sudden, single meander (Gründlingh, 1979). This Natal Pulse (Lutjeharms and
Roberts, 1988) moves downstream with the current at a rate of about 20 km/day.
Features of this kind have also been observed north of the Natal Bight (Gründlingh
and Pearce, 1984; Gründlingh, 1992a), however, all information currently available
suggests that it is only meanders that originate at the Natal Bight that consistently
progress downstream with the current. Theoretical studies (De Ruijter et al., 1999a)
have shown that it is the weak gradient of the shelf at the Natal Bight that will allow
baroclinic instability in the Agulhas Current to occur here and to grow once the core
of the current has been detached from the sharp slope gradient. The trigger for this
meander has been thought to be the adsorption of offshore eddies, the tell-tale signs of
which have been seen in many satellite images in the thermal infrared (e.g.,
Gründlingh, 1986; Lutjeharms and Roberts, 1988). This has recently been proven to
be the case (Schouten et al., 2002). It is interesting to note that some marine animals
such as leatherback sea turtles carefully use all these circulation features to move
about in the ocean (Hughes et al., 1998; Luschi et al., 2003). The question remains
what, if any, effects these unusual meanders have on the shelf waters. This will be
discussed in a section to follow. It is necessary first to describe the wind regimes over
this shelf region.
4.1 Winds along the shelf off south-eastern Africa
Tropical cyclones hardly ever reach this coastline, in contrast to that of Madagascar
and Mozambique (Jury and Pathack, 1991). On the infrequent occasions when they do
arrive at the coast (e.g., Poolman and Terblanche, 1984), one would expect the
24
shallow waters of the shelves over which they move to be thoroughly mixed.
Otherwise the shallow waters, as measured by moored current meters (upper ~20 m),
follow the reigning winds closely (Pearce et al., 1978).
Coastal winds for the region have been analysed in detail by Schumann (1989). He
has shown that the main wind axis is parallel to the coast. At Durban the wind is 5
times more likely to blow along the shelf than across it, whereas at East London (viz.
Fig. 20-011) it is three times more likely. Average wind speeds are about 2.5 m/s at
Durban; 3.2 m/s at East London (for 1984). The average wind speeds along the coast
and across it were not very different. The north-easterly wind and the south-westerly
winds both occur about 50% of the time (Schumann and Martin, 1991), with both
showing seasonality in wind speed, the north-easterly winds having slightly greater
seasonality. During summer the alongshore component of the wind is considerably
higher (Hunter, 1988), particularly farther downstream.
An important additional wind process for the coastal waters is diurnal land and sea
breezes. These can exhibit speeds of the same magnitude as those brought about by
normal synoptic systems (Hunter, 1988). Hunter (1981) has used offshore wind
observations to show that land breezes can here extend at least 60 km seaward. The
direction of these winds may have a decided influence on cloud formation,
precipitation over the shelf as well as coastal runoff.
As mentioned before, it has been demonstrated that cumulus cloud lines frequently
form over the northern Agulhas Current (Lutjeharms et al., 1986b; Lee-Thorp et al.,
1998) but mostly when the winds are along-current, from the north-east. During such
along-current air motion there is an enormous uptake of moisture from the current
(Lee-Thorp et al., 1999; Rouault et al., 2000). About 5 times as much water vapour is
transferred to the atmosphere above the current itself than from ambient waters.
During on-shore winds this moisture is advected inland and may contribute
significantly to moisture convergence and rainfall over the interior of South Africa. In
fact, it has been shown that this leads to local intensification of storm systems and the
concurrent flood events (Rouault et al., 2002). To what extent this leads to measurable
dilutions of coastal waters by river runoff is not known. What is known is that the
presence of the Agulhas Current has a consistent effect on coastal rainfall all along its
Fig. 20-012 The influence on coastal rainfall of the distance of the core of the
Agulhas Current from the coastline. (After Jury et al., 1993.) The abscissa gives
the distance upstream from Port Elizabeth (see Fig. 20-011). The solid curve
gives the distance from the coast to the core of the Agulhas Current as expressed
25
by sea surface temperatures. Note that the distance is greater off the Agulhas
Bank (0 km) and at the Natal Bight (800 km). The broken line shows the coastal
rainfall. Both curves are expressed as standardized departures; that of the
distance from the coast having been inverted for comparison.
northern part (Jury et al., 1993; Fig. 20-012). Wherever the current is close to the
coast, such as between Durban and Port Elizabeth, the rainfall is enhanced; wherever
the current axis diverges from the coastline, such as at the Natal Bight and at the
southern part of the Agulhas Current, coastal rainfall is significantly reduced. This is
not the only process that makes the Natal Bight an unusual shelf region.
4.2 The Natal Bight
The Natal Bight is formed by a landward offset between Richard's Bay and
Durban in an otherwise rather linear coastline (Fig. 20-011). The northern part of the
bight is shallower than 50 m; the southern part deeper. There are some well-
developed canyons in the bathymetry of the continental slope, but these do not extend
onto the shelf, where they have been filled in by sediment (Martin and Flemming,
1988). The major depocentre of the region is the offshelf Tugela Cone (viz. Fig. 20-
013), evidence that the Tugela River is the major source of sediment for this shelf
region. Sediments over the shelf itself consist largely of sand. The percentage is in
excess of 75% over all parts of the Natal Bight shelf except seaward of the Tugela
River (Flemming and Hay, 1988) where mud is the dominant sediment type. Gravel
patches are found largely, but not exclusively, at the shelf edge where scouring from
the Agulhas Current is to be expected. The distribution of sediments (Flemming and
Hay, 1988) is particularly instructive here since it gives a clear indication of the
integrated movement of the bottom waters where other data may not be available.
Fig. 20-013 A conceptual model of the bedload movement on the continental
shelves adjacent to the northern Agulhas Current. (After Flemming and Hay,
1988.)
26
The shelf directly equatorward of the Natal Bight has, for instance, seen hardly any
hydrographic investigations. However, the general bedload dispersal model suggests
that the shelf waters move equatorward here, in clear disagreement with the concept
of a straightforward Mozambique-Agulhas Current continuity. A bedload parting is
found at about 28 °S (Fig. 20-013). Analyses of ships' drift (Harris, 1978) indicate
that poleward of this point the currents over the shelf follow the Agulhas Current 75%
of the time. This implied movement is consistent along the whole coastline except just
downstream of Durban where it again is equatorward. This latter discrepancy may be
due to an embedded lee eddy that is a recurring feature of the circulation at this
location just south of the Natal Bight (e.g., Anderson et al., 1988; Meyer et al., 2002)
and will be discussed in greater detail below.
The submarine bedform distribution is even more instructive. Active submarine
dune fields, moving with the current, are found north of the Natal Bight and along the
shelf break of the northernmost half of the bight. Along the southern half of the shelf
break there is no evidence of the influence of the current, suggesting that it overshoots
here (viz. Fig. 20-011), maintaining some distance from the shelf edge and thus not
affecting the sediments. It is intriguing that off Durban the movement of the mobile
dune field is, by contrast, northward, substantiating the persistence of the lee eddy
surmised to occur here (Pearce et al., 1978; Meyer et al., 2002). Flemming and Hay
(1988) have inferred a complex shelf circulation from the sediment distributions and
the bedforms, consisting of a cyclonic movement over the northern, shallower part of
the bight and a dipolar structure of an inner anticyclonic and an outer cyclonic eddy
over the southern, deeper parts. What is in fact known about the circulation here?
First, the presence of a persistent upwelling cell at the upstream end of the Natal
Bight is the most prominent part of the hydrodynamics of the shelf waters of the Natal
Bight and a fundamental key to understanding the ecosystem of this shelf sea. From
all other perspectives it may be considered to be a semi-enclosed system. The strong
and ever-present Agulhas Current at the shelf edge forms a formidable barrier to
exchanges of water and biota with the open ocean. At the northern end of the bight,
between Richard's Bay and Cape St Lucia, the shelf widens as the current sweeps
poleward. This bathymetric arrangement is believed to lead to topographically
induced upwelling (Gill and Schumann, 1979), as it does elsewhere along the
trajectory of the Agulhas Current.
In this general region sea surface temperatures are about 26 °C in the summer
months, peaking in February (Pearce, 1978) and dropping to about 21 °C in August.
Observations of sea surface temperature in the region (Gründlingh, 1974; Gründlingh
and Pearce, 1990) have shown that off Richard's Bay the temperatures are always a
few degrees lower. As could be expected, the water here is largely Tropical Surface
Water with only the occasional presence of Subtropical Surface Water (Pearce, 1978).
Others (Lutjeharms et al., 2000c) have shown that the purest Subtropical Surface
Waters is found on the shelf edge off Richard's Bay and St Lucia. This sporadic
presence of Subtropical Surface Water on the shelf, otherwise found at depths of 150
m or more offshore (Pearce, 1977b), is highly suggestive. Subsequent investigations
using satellite images (Lutjeharms et al., 1989b) and a dedicated hydrographic cruise
(Lutjeharms et al., 2000c; Meyer et al., 2002) have demonstrated unequivocally that
this is indeed a persistent upwelling cell.
Lower temperatures are observed here more or less continuously, although the
areal extent of the surface expression may vary considerably. This surface expression
seems to have no clear seasonal pattern, neither is it clearly related to potential
27
upwelling inducing winds (Lutjeharms et al., 1989b). Pearce (1978) has shown that
evidence of 16 °C water (Subtropical Surface Water, viz. 20.3) at depths of 125 m,
sometimes less than 100 m depth, on the shelf is intermittent, with no clear pattern. It
can therefore be accepted that this upwelling is not wind-driven. The effect of this
upwelling can be observed at the sea surface along the inner edge of the Agulhas
Current as far downstream as Durban as the colder surface water is dragged
southward as a cool filament (Lutjeharms et al., 1989b). Further evidence for the
nature of this upwelling cell comes from nutrient distributions (Carter and d'Aubrey,
1988; Meyer et al., 2002; Fig. 20-014). It shows that the influence of the upwelling
cell extends over a sizeable part of the bight. Carter and d'Aubrey (1988) have given
historical nutrient values all over the bight and state that there is no clear seasonal
pattern in the occurrence of nutrients. Vertical sections show clearly (Lutjeharms et
al., 2000c) that this nutrient-rich water is upwelled at the St Lucia upwelling cell and
from there moves over the floor of the bight southwards. Further evidence for the
effect of this upwelling cell comes from biological observations.
Fig. 20-014 Distribution of dissolved nitrate at 10 m depth over the Natal Bight
during July 1989. (After Meyer et al., 2002.) Dots represent station positions.
28
The shelf shallower than 200 m has been shaded. The presence of an active
upwelling cell equatorward of Richard's Bay is evident.
The distribution of chlorophyll-a exhibits a very similar pattern to that of the
nutrients (e.g., Meyer et al., 2002), with the enhanced values slightly lower very close
to the coast, implying an active upwelling process taking place during the
observations. Oliff (1973; as quoted by Carter and Schleyer, 1988) has shown how the
phytoplankton production reacts almost instantaneously to an upwelling event at
Richard's Bay. Reviews of the plankton, zooplankton as well as the benthic species
found in the Natal Bight have been given by Carter and Schleyer (1988) and by
McClurg (1988) respectively. These studies were based on information that was
geographically very inhomogeneous, since they had to depend on an eclectic set of
previous collections not designed uniformly to cover the shelf as a whole. From these
scattered observations it is impossible to infer the extent of the biological influence of
the St Lucia upwelling cell over the Natal Bight shelf, particularly over the southern
part.
The waters of the poleward part of the Natal Bight are by contrast unspectacular
and fit well into the ranges, both physical and biological, to be expected at these
latitudes. The nutrients are low (Carter and d'Aubrey, 1988; Meyer et al., 2002),
coming from Tropical Surface Waters. The nutrient concentrations are 1.01 1.86
mol/ (nitrate), 0.48 0.72 mol/ (phosphate) and 3.50 4.69 mol/ (silicate) at
10 m depth. The values on this part of the shelf do not exhibit great differences from
those found in the surface waters of the Agulhas Current. The exception is to be found
close to the Tugela River mouth (viz. Fig. 20-011) where values of all nutrients are
higher during floods and chlorophyll-a values are enhanced (Carter and Schleyer,
1988). This is reflected in greater densities of fish larvae at such times (Beckley and
van Ballegooyen, 1992). When this outflow was directly observed during such a flood
event, the indications were that most of the outflow occurred at a depth of 30 m and
extended at least 25 km offshore. Salinities and temperatures over the shelf otherwise
closely follow those of the Agulhas Current itself, also its seasonal cycle.
The circulation in the southern part of the bight is much harder to establish.
Remote sensing has suggested a cyclonic eddy (Malan and Schumann, 1979) and this
has been considered the main element in many conceptual portrayals ever since (e.g.,
Pearce, 1977b, Gründlingh and Pearce, 1990; Schumann, 1987; Harris, 1978).
Observations show (Pearce, 1977b) that close to the coast the currents only follow the
Agulhas Current 50% of the time. Ship's drift close inshore (1.6 km) is also about
equally divided between poleward and equatorward drift (Harris, 1964; Pearce et al.,
1978), agreeing with wind frequencies. The closer to the current, the greater the
tendency is to follow its direction closely (Harris, 1978). The only quasi-synoptic
hydrographic survey of the bight as a whole (Lutjeharms et al., 2000c) gives no
indication of a consistent circulation. There are indications that the location of the
edge of the Agulhas Current may show greater shifts in location along the edge of this
particular part of the shelf than farther up- or downstream (Gründlingh and Pearce,
1990) and that shear edge eddies may play an important role on occasions (e.g.,
Lutjeharms and Roberts, 1988; their Fig. 10b). Putting it all together, Pearce et al.
(1978) have concluded, on the basis of an eclectic set of observation, that "at any one
time a succession of eddies of a variety of scales (are) generated by shear processes or
meteorological forcing probably exists in the (Natal Bight)" and this is as good a
29
summary of what is currently known as data will allow. Directly south of the bight,
adjacent to the city of Durban (Fig. 20-011), the situation seems much simpler.
As mentioned above, a number of investigators (e.g., Pearce et al., 1978;
Schumann, 1982; Anderson et al., 1988; Lutjeharms et al., 2000c and Meyer et al.,
2002) have pointed out the presence of a cyclonic eddy directly off Durban, in the lee
of the broader shelf that forms the Natal Bight. Currents measured off Durban do
show a dominant north-eastward component (Fig. 20-015). Ship's drift close inshore
(1.6 km) is similar to further north on the shelf about equally divided between
poleward and equatorward drift (Harris, 1964; Pearce et al., 1978), which agrees with
wind frequencies. Drifters have shown the same bi-polar tendency. Results presented
by Tripp (1967) are also in agreement, showing that the current sets north-eastwards
only about 50% of the time, with speeds of between 0.25 to 0.51 m/s. Current
observations for a period of a month (Schumann, 1988b), as well as over shorter
periods (Gründlingh and Pearce, 1984), indicate frequent current reversals here. There
is therefore abundant evidence that this eddy does not seem to be present all the time.
Current reversals at its location are measured over the full depth, indicating the
Fig. 20-015 Current observations off Durban and Port Edward. (According to
Schumann, 1982.) For the location of these two places, see Fig. 20-011. The large
dot indicates the location of the observations; the small dots the tips of current
arrows from the station location. These arrow points show speed and direction
integrated over the top 100 m of the water column. Each dot represents one spot
measurement.
barotropicity of this eddy. The main driving force of the shelf circulation at this
location is most probably the Agulhas Current and not the wind (Schumann, 1981). It
is clear from hydrographic observations that there is considerable upwelling in this
eddy, bringing nutrient rich water closer to the surface, but this also occurs in a very
spasmodic way.
30
When there is no eddy present there may be a mixed layer to a depth of 50 m with
a maximum gradient in nitrate-nitrogen lying below 100 m. With an eddy present this
nutricline can be lifted to about 40 m (Carter and d'Aubrey, 1988) with a concurrent
drop in surface temperatures. This surface cooling has also been observed in satellite
thermal infrared observations (Lutjeharms and Connell, 1989). Average values of
nitrates in the Durban eddy have been measured around 3.33 mol/, but maximum
values have reached 16.79 mol/, demonstrating the effect of the eddy's presence at
times. These higher values in nutrients do not seem to be reflected in higher
chlorophyll-a values. There is evidence of higher zooplankton biomass here, but only
intermittently. Phytoplankton production rates are below 1 gC/m2/ day (Burchall,
1968). There has been a greater density of fish larvae off Durban on occasion
(Beckley and van Ballegooyen, 1992), but not consistently (Beckley and Hewitson,
1994). A hydrographic cruise covering the whole Natal Bight has clearly
demonstrated (Lutjeharms et al., 2000c; Meyer et al., 2002) the vertical structure of
this eddy compared to the waters over the rest of the shelf. The upwelling of nutrient-
rich water did not enhance chlorophyll-a values on that occasion either.
In summary, a cyclonic lee eddy is a recurrent, but not ever-present part of the
circulation on the shelf directly off Durban, but seems to have hardly any local
biological impact, perhaps because it is not enduring. There is evidence that this
cyclonic eddy and its nutrient contents form the core to incipient Natal Pulses
(Lutjeharms and Roberts, 1988).
When a Natal Pulse forms, due to an offshore eddy or otherwise, it seems to carry
the Durban lee eddy with it (Lutjeharms et al., 2003b) all the way to the southern tip
of the continent, intensifying the cyclonic motion in the eddy in its downstream
journey. This would imply that inshore currents from Durban downstream would
experience a sudden, but short-lived, reversal. As mentioned before, such a reversal
has actually been observed (Lutjeharms and Connell, 1989) and directly related to a
passing Natal Pulse. This incorporation of the Durban eddy in Natal Pulses also
means that a part of the shelf fauna of the Natal Bight will intermittently be carried
away. The effect of such a mechanism on the shelf biota is not known. Most local fish
seem to spawn in Cape waters downstream and migrate upstream in July and
November (Garratt, 1988). Only geelbek (Atractoscion aequidens) and seventy-four
(Polysteganus undulosis) appear to spawn off the east coast of southern Africa. A
small proportion of the fish caught off Natal are present throughout the year, most are
distinctly seasonal, being either summer or winter species (Van der Elst, 1988). The
most dramatic of the upstream migrations is that of the pilchard Sardinops ocellata,
that takes place close inshore and is therefore very noticeable. No correlation with the
passing of Natal Pulses in any of these fish migrations has to date been established.
4.3 Between Durban and the Agulhas Bank
Downstream of Durban the bathymetry changes dramatically (Fig. 20-011). Here the
shelf is narrow and without any indentations to speak of. This configuration of the
continental border has a controlling influence on the behaviour of the Agulhas
Current. The current closely follows the edge of the shelf (Gründlingh, 1983) and the
currents on the shelf move largely in sympathy (Fig. 20-015). Current speeds up to 1
m/s have been measured to 10 km off the coast. The currents at the shelf edge are, on
average, about 0.5 m/s just downstream of Durban (based on ships' drift; Tripp,
1967), but increase steadily till they are 2.16 m/s off Port Elizabeth. Current speeds
31
inshore are much lower and vary in a seemingly random manner from 0.38 to 0.77
m/s. At Port St Johns there is a minor coastal offset (viz. Fig. 20-011) and this seems
to cause a higher frequency of counter currents (Harris, 1964), up to 40% of the time
(Tripp, 1967). Observed currents at Port Edward (Fig. 20-015) are with the Agulhas
Current, but somewhat reduced in speed. The strong currents are reflected in the sand
transport and bedform patterns along this whole shelf edge. Substantial underwater
sand dunes are formed (Flemming, 1978, 1980, 1981) and this whole dune field is
mobilised to move downstream with the current. It has been surmised that at certain
locations where canyons cut into the shelf they act as sediment traps, carrying
sediment off the shelf.
Surface drifts over this shelf region, as measured by drifters (Anderson et al.,
1988), are parallel to the current, but in both alongshore directions, suggesting that
they are driven largely by the wind. Movement at greater depths is with the current
(Schumann, 1982; 1987) except during very strong south-westerly winds when the
shelf currents may briefly change direction. The latter is also reflected in observations
of ships' drift (Pearce et al., 1978), although the incidence of current reversal is much
reduced here compared to the movement in the Natal Bight, demonstrating the greater
influence of the juxtapositioned Agulhas Current. From a direct comparison of current
meter data and concurrent wind records it has been shown (Schumann, 1981) that the
correlation between these signals is smaller than expected. Many current reversals
therefore seem likely to be due to the passing of a Natal Pulse. However, there are
other possible low frequency fluctuations in the currents on the shelf on this coast.
Schumann (1981; with Perrins, 1982) has analysed current records for signs of
tidal and inertial signals. He found limited inertial motion; the energy spectrum being
dominated by much longer periods that were assumed to be associated with variations
in the Agulhas Current. Schumann (1986), in an investigation of the bottom boundary
layer along this coastline, has furthermore demonstrated that in 50 m of shelf water
Ekman veering took place over the lower 35 m. This would cause some upwelling of
deeper water, possibly enhancing biological productivity.
Observations of the vertical distribution of nutrients do indeed show higher
nutrient levels at depth (Carter and d'Aubrey, 1988) and over the deeper parts of the
shelf. In general shelf waters were poor in nutrients (nitrate: 2.48 mol/ mean; 15.80
mol/ maximum) as is to be expected where surface waters of the Agulhas Current
dominate. No observations have to date been made during the passage of a Natal
Pulse and its imbedded Durban eddy. Conceivably such an event could cause a
sudden, but short-lived elevation of nutrient concentrations. The effect of such pulses
on the productivity along this stretch of shelf remains unknown. Observations of
zooplankton (Carter, 1977) indicate higher biomass at the edge of the Agulhas
Current here than upstream. Ichthyoplankton observations (Beckley and van
Ballegooyen, 1992) reveal the same, but with great variability in space and time.
To recapitulate: the influence of the northern Agulhas Current on the shelf
circulation seems to start only at about 28° S. The circulation in the wider shelf region
of the Natal Bight is principally influenced by the passing current in the creation of an
upwelling cell at St Lucia. By inserting nutrients into shelf waters this cell may have a
controlling influence on the ecology of the whole Natal Bight. At the southern end of
the bight the current drives a persistent lee eddy off Durban. The movement of waters
on the narrow shelf south of Durban are for the most part parallel to the Agulhas
Current. With the possible exception of what happens during the passage of a Natal
Pulse, the circulation on this particular shelf region consequently seems
32
uncomplicated, with low productivity and biological activity. The exception seems to
be at Port Alfred, at the eastern tip of the Agulhas Bank.
5. Southern Agulhas regime
The Agulhas Bank forms the triangular continental shelf south of Africa (Fig. 20-
016). It lies between Port Elizabeth in the east and Cape Town in the west. It is about
250 km at its widest. A shallow part, the Alphard Rise, where the shelf is widest (viz.
Fig. 20-016), constitutes the border between the western and the eastern Agulhas
Bank that, as a result of this partition, have distinctly different characteristics. This is
exemplified by the contrasting sedimentary nature of the eastern and western Agulhas
Bank.
The sediments on the eastern bank are dominated by shelly fragments whereas
there are mainly foraminiferal oozes on the western bank (Rogers and Bremner,
1991). The organic matter content of the eastern Agulhas Bank sediments lies
between 0.0 and 3.9 % (per unit mass) compared to 4.0 to 11.9 % on the western
Agulhas Bank This places the western bank squarely in the province of the Benguela
upwelling system
Fig. 20-016 Bathymetry of the continental shelf off the south coast of South
Africa in km (after Simpson, 1974; Dingle et al., 1987a) and its major circulatory
elements. This covers the southern Agulhas Current regime. Areas shallower
than 1 km are shaded. The broken line denotes the edge of the continental shelf
as defined by the 200 m isobath. Upwelling is shown by hatching. Place and
feature names used in the text are given here.
(Dingle et al., 1987b) A further reason for this difference is the contrasting
characteristics of the offshelf currents on either side.
On the eastern side the warm waters of the intense southern Agulhas Current
follow the shelf edge (Fig. 20-016), but much less closely than upstream. The current
33
starts to meander near Port Elizabeth and these meanders grow in amplitude
downstream (Lutjeharms et al., 1989a). The meanders come with cyclonic shear edge
eddies and attendant warm plumes. These plumes in general are shallow features, but
may eventually spread their warm water over large parts of the adjacent shelf. By
contrast, on the western side of the Agulhas Bank the sluggish Benguela Current
carries cold water equatorward, but this is complicated by the presence of a lee eddy
directly to the east of the Agulhas Bank (Penven et al., 2001a), the recurrent passage
of Agulhas rings (Lutjeharms and Valentine, 1988; Boebel et al., 2003) and the
intermittent advection of warm Agulhas surface water in the form of Agulhas
filaments (Lutjeharms and Cooper, 1996). However, this shelf region is wide and the
inner parts may be less affected by the offshore currents than by winds.
5.1 Wind regimes south of Africa
As is the case along the east coast, the winds along the south coast are mainly parallel
to the coastline, more so at Port Elizabeth than in the central part of the south coast
(Schumann, 1989). The ratio of offshore/onshore to coast-parallel winds is also lower
along the south coast to that of Port Elizabeth where the winds are more strongly
oriented parallel to the coast. North-easterly winds show the greatest seasonality with
a greater than 40% occurrence frequency in austral summer, dropping to 25% in
winter (Schumann and Martin, 1991). Average speeds for north-easterly winds at Port
Elizabeth are greater than 4 m/s in summer, but only 1.5 m/s in winter. South-westerly
winds show an inverse seasonal occurrence compared to north-easterly winds, but the
wind speeds always lie around 4 m/s. Based on ships' reports, Jury (1994) has shown
that on the eastern Agulhas Bank the most frequent and the strongest winds are from
the east in summer and from the west in winter. Winds are significantly weaker over
the southern parts of the shelf compared to closer to the coast. These winds and their
directions naturally are part and parcel of the global ENSO and other perturbations
(e.g., Schumann, 1992). Little work has to date been carried out to pin down these
relationships.
A significant proportion of these winds come about as a consequence of coastal
lows. These are formed as a result of the topography of southern Africa (Gill, 1977),
most often associated with cold fronts that move from west to east (Hunter, 1987).
The coastal lows move along the south coast with periods of 2 to 5 days and
propagation speeds of 14 to 20 m/s. Analyses of the climatological variability of the
major axis winds along the coast of the Agulhas Bank (Schumann et al., 1991) has
therefore shown a distinct spectral peak at 6 days.
Over the western Agulhas Bank the wind patterns are significantly different to
those on the eastern bank. At Cape Town the most seasonal wind is the south-easterly
wind with an 80% occurrence frequency in summer and a 40% frequency in winter.
Average speeds in summer are in excess of 5 m/s; only 1.5 m/s in winter. By contrast
the north-westerly winds are only slightly more prevalent in winter and maintain
average speeds of about 3 m/s.
Wind strength is increased over the Agulhas Current itself. The loss of heat from
the Agulhas Current to the atmosphere is about 200 W/m2 higher than that of ambient
water masses (Rouault et al., 2000). During along-current winds an atmospheric
moisture and thermal front develops at the inshore edge of the current off Port Alfred,
where the current passes a distinct upwelling cell.
34
5.2 The Port Alfred upwelling cell
As mentioned before, if the theoretical portrayal of Gill and Schumann (1979) holds,
wherever a western boundary current moves from a narrow shelf past a wider shelf a
degree of upwelling should be experienced. This has been observed south of
Madagascar, at the northern corner of the Natal Bight and even in the Kuroshio
system (Lutjeharms et al., 1993). It would therefore be expected to occur also where
the Agulhas Current starts flowing along the Agulhas Bank and there it has indeed
been observed (Lutjeharms et al., 2000a).
Thermal infra red imagery from satellite shows that this upwelling has its centre
off Port Alfred (viz. Fig. 20-016), but that cold water inshore of the Agulhas Current
may extend up to a maximum of 300 km upstream. On average it is about 30 km wide
and extends 180 km along the edge of the current (Fig. 20-017). Water masses in the
upper 200 m of the Agulhas Current are Subtropical Surface Water with the
occasional pulse of Tropical Surface Water, mostly at the landward side (Gordon et
al., 1987). During those times that the upwelling is evident at the sea surface the water
in the upwelling cell is all South Indian Central Water (Lutjeharms et al., 2000a). This
implies that it is upwelled onto the shelf from offshore waters deeper than 400 m and
this has been confirmed by hydrographic observations in the region (Goshen and
Schumann, 1988; their Fig. 8). Nutrient values in the upwelling cell can exceed, for
example, 20 mol/
Fig. 20-017 An ensemble of outlines of cold water of the Port Alfred upwelling
cell inshore of the Agulhas Current, using the 17 °C isotherm as an indicator.
(As given by Lutjeharms et al., 2000a.) The 200 m isobath is shown as a dotted
line and the shelf inshore of this isobath shaded.
35
at 100 m, whereas they are less than 5 mol/ on the adjacent shelf. This upwelling
cell may furthermore have considerable implications for the Agulhas Bank as a
whole.
First, it forms one of what may be considered to be the three main
physical/chemical provinces of the bank. The others are the greater part of the shelf
and the coastal upwelling regime of the western Agulhas Bank (Lutjeharms et al.,
1996). These provinces are well-defined, considering the poor and inhomogeneous
distribution of hydrographic stations available. Second, from this upwelling cell water
colder than 10 °C moves over the bottom of the Agulhas Bank (Lutjeharms and
Meyer, 2004). The seasonal thermoclines over the bank are unusually intense. It is
believed that this is partially due to the continual input of cold water along the
bottom. It has previously been surmised that this cold water is upwelled inshore of the
Agulhas Current along the eastern edge of the bank (Chapman and Largier, 1989). To
date little direct evidence for this has been found. In fact, almost all hydrographic
sections across the bank show the water colder than 10 °C near the shelf edge only in
the vicinity of Port Alfred. If this upwelling cell plays such a crucial role in the
stratification of the whole Agulhas Bank, how permanent is it?
Regrettably, there are no data to address this important question. Surface
observations show that it is present almost 50% of the time (Lutjeharms et al., 2000a),
but this surface outcropping may be largely wind dependent, as has been observed at
sea (Rouault et al., 1995). With the limited hydrographic data available, the presence
of upwelled water at depth seems considerably more enduring. If this upwelling cell is
so persistent, how does it affect the distribution of primary productivity and biota at
higher trophic levels?
A chlorophyll-a maximum zone extends from the vicinity of Port Alfred roughly
along the 100 m isobath across the eastern Agulhas Bank (Probyn et al., 1994). This
"upwelling ridge" has primary production rates of 104 mg/m2/h, whereas the shelf
near Port Alfred boasts values of 888 mg/m2/h. Values at the shelf break are 231
mg/m2/h. In this respect it is interesting that ocean colour observations from satellite
(Lutjeharms and Walters, 1985) show slightly enhanced values along the whole
landward edge of the Agulhas Current. By contrast the zooplankton biomass is low
near Port Alfred (Verheye et al., 1994), increasing westward. Spawning of chokka
squid, as detected by eggs trawled, show a distinct concentration at Port Alfred, with
a downstream decrease across the shelf, downstream (Augustyn et al., 1994). About
predators much less is known (Smale et al., 1994). The size distribution of fish varies
noticeably across the Agulhas Bank (Japp et al., 1994), the larger fish being found on
the eastern Agulhas Bank, some species densities higher at the horizontal thermal
gradients associated with the shelf edge (Barange, 1994). The biological influence of
the Port Alfred upwelling cell therefore seems indirect, particularly via the inflow of
bottom water over the eastern Agulhas Bank.
5.3 The eastern Agulhas Bank
Much has been made of the ridge of cold water (< 10 °C) that overlies the 100 m
isobath over much of the eastern Agulhas Bank (e.g., Boyd and Shillington, 1994;
Verheye et al., 1994; Probyn et al., 1994). This ridge is evident in temperature
sections across the bank; at times it crops out, producing areas of cold water at the sea
surface (e.g., Walker, 1986; Swart and Largier, 1987) over the bank. The location of
this ridge corresponds with the distribution of a number of copepod development
36
stages (Largier and Swart, 1987), medium size chokka squid (Augustyn et al., 1994)
and a subsurface chlorophyll-a maximum at the nutricline (Probyn et al., 1994). It has
been proposed (Lutjeharms and Meyer, 2004) that this ridge of cold, nutrient-rich
water originates in
Fig. 20-018 Time series of the temperature profile for a hydrographic station
on the central Agulhas Bank for the period 1974 to 1979. Dots indicate standard
depths at which nutrient samples were taken. (After Lutjeharms et al., 1996.)
the Port Alfred upwelling cell. It enhances the thermal stratification of the water
column from below over most of the bank and brings nutrients onto the full extent of
the shelf. These nutrients may not be available to the phytoplankton throughout the
year, but follow an annual cycle. Observations show (Carter et al., 1987) that the
subsurface chlorophyll-a peak is closely associated with the behaviour of the seasonal
thermocline. This stratification on the Agulhas Bank follows a rather peculiar cycle
(Eagle and Orren, 1985; Lutjeharms et al., 1996; Fig. 20-018).
During the winter months storm winds and convection mix the upper water column
to depths of 75 m or more; in summer fewer extreme wind events and increased
insolation establish a seasonal thermocline that may be at 50 m or deeper. Summer
winds have been shown to be too weak (Largier and Swart, 1987) to break down the
seasonal thermocline. The winter mixing of the water column raises the temperature
of the bottom water in winter, being warmest in July (Swart and Largier, 1987). All
this is normal for shelf regimes in the subtropics. On the Agulhas Bank this seasonal
stratification is however enhanced by cold water from below (Fig. 20-018) thus
establishing a seasonal thermocline and nutricline about twice as strong as would
normally be expected. It is believed that the inflow of all this cold, nutrient-rich water
is from the Port Alfred upwelling cell.
The result of this seasonal behaviour of the water column is also reflected in the
temperature/salinity characteristics of the Agulhas Bank. In spring the temperature
range is restricted to 14 18 °C, salinity to 35.0 to 35.5; by contrast, in winter this
ranges from 10 23 °C, salinity 34.6 to 35.8. The nutrients follow suite. Average sea
surface temperatures are between 21 26 °C over the eastern Agulhas Bank,
representing the horizontal offshore gradient towards the Agulhas Current (Schumann
37
and Beekman, 1984). In winter this is reduced to 16 19 °C. In summer the average
temperature difference between the water at the surface and at 80 m depth is as high
as 11 °C; in winter the highest gradient is 5 °C. With the seasonal collapse of the
nutricline the subsurface chlorophyll-a peak also disappears (McMurray et al., 1993).
Nevertheless the seasonal thermocline on the eastern bank is usually shallower and
better developed compared to that of the western part of the bank.
The movement of the waters of the eastern Agulhas Bank can most probably be
placed into three categories: the very surface layer, the bottom layer and the rest of
the water column. The very surface layer moves largely with the wind (Lutjeharms et
al., 1986a). Comparison of the movement of a drifter with progressive wind stress
vectors rotated 35° to the left has demonstrated this quite admirably (Harris, 1978).
Analyses of ships' drift, corrected for windage on ships' superstructure, show (Tripp,
1967) no distinct patterns or seasonal cycles. Since the dominant wind directions are
parallel to the coast, so will the top water layer therefore move. This is particularly
important from a pollution point of view since tar balls (Shannon and Chapman,
1983; Shannon et al., 1983) will be beached only under onshore wind conditions. The
bottom waters, where they have been measured (Swart and Largier, 1987), move
largely parallel to the isobaths and in the direction with the Agulhas Current. The
movement of the rest of the water column is more complex. Schumann and Perrins
(1982) have shown that the highest energies on the shelf are to be found at inertial
periods. The bottom motion described earlier consists of the residual drift after the
inertial current signals have been removed. The influence of the Agulhas Current is
not always clear.
As mentioned above, the major characteristic of the Agulhas Current along the
shelf edge of the eastern Agulhas Bank is the presence of shear edge eddies of various
sizes (Lutjeharms et al., 1989a). These eddies move downstream with the current and
are most prevalent in the concave part of the shelf edge (Lutjeharms et al., 2003a).
There is evidence (Hutchings, 1994) that the spawning of anchovy takes place
preferentially just inshore of the shelf edge where these eddies are common. The
passage (Schumann and van Heerden, 1988) of such eddies inverts the shelf edge
current that is usually dominated by the Agulhas Current. Cold water is upwelled in
the core of these cyclonic features and is has therefore been surmised that they are
instrumental in bringing this cold water onto the shelf (Chapman and Largier, 1989).
This has been observed very infrequently (e.g., Schumann and Beekman, 1984). The
warm water plumes associated with the shear edge features bring warm Agulhas
Current water onto the shelf and this may enhance the shelf thermocline. In general
these plumes may represent a relatively thin layer of warm water (e.g., Goschen and
Schumann, 1990) and their subsequent movement over the shelf may be almost
entirely driven by winds (Goschen and Schumann, 1994).
Once past the southern tip of the Agulhas Bank, the Agulhas Current proper moves
offshore, but some of the plumes may be drawn along the edge of the western
Agulhas Bank as Agulhas filaments.
5.4 The western Agulhas Bank
Agulhas filaments are relatively shallow features (~ 50 m) and may carry warm
surface water as far as the coastal upwelling zone off south-western Africa
(Lutjeharms and Cooper, 1996). To date there has been only occasional evidence
(e.g., Mitchell-Innes et al., 1999) that warm surface water from these filaments may
38
intrude over the shelf in an analogous fashion to that of plumes adjacent to the eastern
Agulhas Bank. Why Agulhas filaments are drawn so rapidly equatorward is also not
entirely clear. This has been assumed (Lutjeharms and Valentine, 1988) to be due to
passing Agulhas rings. Such ring prevalence may be the origin of the hypothesised
shelf edge jet (e.g., Hutchings, 1994) that has been put forward as a mechanism to
move anchovy larvae northward towards the coastal upwelling region of the west
coast (e.g., Boyd et al., 1992). Then again, whenever direct current observations of
the currents over the shelf edge have been made (e.g., Fowler and Boyd, 1998) no
evidence for any persistent north-westerly setting current could be found.
However, there is strong evidence that there is a tendency for the circulation off
the shelf of the western Agulhas Bank to be cyclonic, which would lead to currents at
the shelf edge to move poleward instead. Modelling (Lutjeharms et al., 2003a) and
hydrographic observations (Penven et al., 2001a) now suggest that this is due to a
cyclonic eddy driven by the Agulhas Current in the lee of the Agulhas Bank. The
discrepancy between an equatorward shelf edge jet and a poleward movement driven
by an Agulhas Bank lee eddy had not yet been resolved. Apart from the difference in
current behaviour at the shelf edge, the western Agulhas Bank also differs from the
eastern bank by extensive coastal upwelling.
The main, wind-driven upwelling along the west coast normally starts at Cape
Point (south of Cape Town, viz. Fig. 20-016), but during about 10% of the time there
also is contiguous upwelling all the way between Cape Agulhas and Cape Point
(Lutjeharms and Meeuwis, 1987; Lutjeharms and Stockton, 1991). This latter
upwelling, as expressed as a biologically productive region, may extend from less
than 20 to 100 km offshore (Mitchell-Innes et al., 1999). At Hermanus and Gansbaai,
between Cape Point and Cape Agulhas, annual surface temperatures range (Largier et
al., 1992) from 17 °C in January to 15 °C in August with a small maximum in May.
With a higher incidence of south-westerly winds in summer, the upwelling here is
most intense in that season (Boyd et al., 1985), as can easily be modelled (Penven et
al., 2001b). Constant westerly or south-westerly winds may induce downwelling, but
upwelling can recur within a matter of days under the influence of winds of the
correct direction and sufficient speed (Jury, 1988). The vertical temperature gradients
in this upwelling are strongest in summer, weakest in winter (Schumann and
Beekman, 1984). Cool, low-salinity Central Water was upwelled to within 20-40 m of
the sea surface from January to April during a survey in 1975 (Boyd et al., 1985) and
from September to November when south-easterly winds are dominant. Only between
February and April was it observed to break the surface. Climatologically it is also
present in July and December (Lutjeharms and Stockton, 1991). Vertical stratification
was best developed in summer, due to the uplift of cooler Central Water and the
onshore advection of warm surface water from the Agulhas Current and the Agulhas
Bank. This is reminiscent of the cycle of stratification over the central Agulhas Bank
(Eagle and Orren, 1985; Fig. 20-018). Nutrients moved closer to the surface in spring
and summer, causing a modest spring bloom (Mitchell-Innes et al., 1999), but
generally did not penetrate the thermocline until vigorous mixing to a depth of 60-80
m in winter.
Over the rest of the western Agulhas Bank the stratification seems much weaker
than over the eastern bank. The uplift of the isopycnals towards the coast over this
shelf region is naturally evident in the distribution of nutrients with high values
covering the greater part of the shelf (Lutjeharms et al., 1996) than over the eastern
Agulhas Bank.
39
The prevailing currents on the western Agulhas Bank are in a north-westerly
direction in summer; in winter those along the southern parts of this coastline may be
south-eastward (Harris, 1978). Water characteristics are the same above the
thermocline than on the eastern Agulhas Bank, but below the thermocline the waters
are slightly less salty and have lower nutrient loads (Chapman and Largier, 1989).
This has been taken as evidence that this water has an Atlantic rather than an Indian
Ocean origin and that the bottom water here on average moves in a south-easterly
direction. In general the water movement here is parallel to the isobaths (Largier et
al., 1992), but in both directions. Nevertheless, inertial currents may still be the
dominant movements on shorter time scales (Schumann and Perrins, 1982).
Evidently, the coastal upwelling on the western Agulhas Bank will have a dramatic
effect on the ecosystem of this part of the bank. In contrast to the eastern bank,
chlorophyll-a values are considerably higher (Largier et al., 1992). Phytoplankton
concentrations are lower (De Decker, 1973) than in the upwelling region along the
west coast, both measured as numbers of cells and as settled volume. Remarkably,
zooplankton settled volumes are almost consistently higher. Biota in higher trophic
levels (e.g., Roel et al., 1994) do not show any patterns clearly related to the
upwelling, including euphausids.
5.5 Coastal processes
The wind-driven upwelling along the western part of the Agulhas Bank is a specific
coastal process, but there are a number of other such locations and processes on the
Agulhas Bank that are largely divorced from what happens over the broader expanses
of the shelf. Perhaps the most important of these is intermittent upwelling at the
promontories along the coast of the eastern Agulhas Bank (Schumann et al., 1982;
viz. Fig. 20-016).
Fig. 20-019 Sea surface temperatures and wind speeds at Gouriqua (near
Mosselbaai, viz. Fig. 20-016) on the south coast of South Africa during March
1984. Wind speed is shown as a curve; wind direction by letters on the abscissa
of the wind speed curve. (According to Lutjeharms, 1998.) Temperatures rise
and fall at the coastline in direct response to the direction and speed of the local
wind.
40
On the whole the seasonal cycle of sea surface temperatures at this coast is
considerably better developed than that along the northern Agulhas Current. Average
temperatures at Port Elizabeth, for example, range from 21 °C in January to about 15
°C in July/August (Lutjeharms, 1998) whereas those at East London vary only from
19 °C to 17 °C. Intermittent coastal upwelling along the south coast is superimposed
on this seasonal sequence. It occurs only at prominent capes (Schumann et al., 1982)
under suitable wind directions (e.g., Goschen and Schumann, 1995) and with winds
above a certain threshold (Schumann et al., 1995). Upwelling is seen less than 10%
of the time, the highest incidence being in summer. The local effects can be quite
dramatic (Fig. 20-019). Beckley (1983, 1988) has reported temperatures dropping by
8 °C over a period of 24 h at a cape site compared to one less than 40 km away.
Goschen and Schumann (1995) have observed reaction times of a few hours. These
upwelling events may last from one to 5 days (Schumann, 1999). Wind events in
along-coast winds have a frequency peak between 4 and 8.5 days, with a maximum at
7 days (Schumann et al., 1991) indicating the driving role of the winds.
Nevertheless an analysis of chlorophyll-a intensity at this coastline (De Villiers,
1998) suggests that the correlations between primary productivity and sea surface
temperature is much better than between primary productivity and upwelling inducing
winds. Along the very farthest eastern side of the Agulhas Bank it may be difficult to
distinguish between local, wind-induced upwelling and the advection into the region
of cold water that has been upwelled in the Port Alfred upwelling cell (e.g.,
Schumann et al., 1988). Upwelling events of this kind have in fact been observed to
attract large flocks of sea birds. This phenomenon remains unexplained, since the
short duration of most upwelling events would not result in greatly enhanced primary
productivity. It has been hypothesised (Schumann et al., 1988) that the convergence
associated with an upwelling front may have caused concentration of food items for
birds. The counterpart of coastal upwelling of cold water is advection of warm water
into coastal embayments.
Along the far eastern side of the Agulhas Bank the likelihood of an intrusion of
warm surface water from the adjacent Agulhas Current right to the coast may be high.
Under southerly wind conditions water from plumes may reach the coastline and
cause a temperature rise of 3 °C or more in surface water. They may also cause a
distinct layering in offshore water. To some extent the coastal morphology plays a
role in the manner in which offshore events may influence the water masses or
circulation at the coast. At certain locations, such as Algoa Bay at Port Elizabeth (viz.
Fig. 20-016), the local winds may create circulations restricted to a bay (Goschen and
Schumann, 1994). A prominent case of localised circulation is found in False Bay, the
large, rectangular Bay on the most western tip of the western Agulhas Bank, just
south-east of Cape Town (viz. Fig. 20-016).
The surface circulation of this bay is complex (Gründlingh and Largier, 1991). On
the eastern side there is intermittent, wind-driven upwelling (Cram, 1970) that brings
nutrient rich water to the surface (Taljaard, 1991). The border of this upwelling may
on occasion form a zonal front across the mouth of the bay (Lutjeharms et al., 1991).
Only one hydrographic survey of the bay as a whole has been made (Gründlingh,
1992b, Taljaard, 1991) and as part of that investigation two sections across the bay
were repeated shortly after. These repeat measurements have dramatically
demonstrated the variability to be expected.
41
During the observations in 25-27 April 1989, cold, fresher upwelled water from
farther east along the coastline was advected into the western side of the bay, creating
a meridional front, evident throughout the water column. During the repeat survey
between 27 and 29 April 1989, the situation had changed completely. Now there
again was a meridional front near the centre of the bay, but with the cold water on the
east, brought about by substantial upwelling on that side. A shallow thermocline (30
m) remained well-developed in the northern, shallower part, but more variable in the
southern part of the bay. Surface currents are also very variable and show a range of
different patterns (Atkins, 1970), probably wind generated (Jury, 1991). The bay is
surrounded by mountains, creating a strong orographic effect on the air flow. At
depth, by contrast, the currents seem much more steady and consistent (Gründlingh et
al., 1989). Water enters on the western side and leaves on the eastern side. As might
be expected the vertical stratification is very seasonal, particularly in the southern,
deeper parts (Gründlingh, 1992b). Huge demographic shifts to the shores of the bay
have placed enormous pressure on its self-cleaning ability. As in many other parts of
the shelf seas discussed above, ignorance of the flushing rate and many other aspects
of the circulation and the ecosystem make its proper management hazardous.
To sum up: the shelf waters of the Agulhas Bank may be placed into three
physical/chemical provinces according to the influence of the Agulhas Current, or
lack thereof. These are: the Port Alfred upwelling cell, the coastal upwelling of the
western Agulhas Bank and, third, the wider shelf outside the immediate influence of
the Agulhas Current. The Port Alfred upwelling cell may be permanent at depth, with
a surface outcropping driven by local winds. Cold, nutrient-rich water is brought onto
the shelf here and may spread over the greater part of the Agulhas Bank. There is a
high likelihood that this inflow of bottom water is the driving force for the very
intense seasonal thermoclines over the wider shelf region. The northward advection of
warm Agulhas plumes is assumed to play a minor role. The unusual intensity of the
stratification over this shelf may be crucial to the local ecosystem and, in particular, to
the successful spawning of economically important fish species. The upwelling on the
western Agulhas Bank is a seasonal phenomena and forms part and parcel of the
greater Benguela upwelling system.
6. Future directions
From the above brief review it should be abundantly clear that the multidisciplinary
characteristics of this part of the world ocean are very imperfectly understood. This is
particularly true for the coastal oceans. This ignorance is easily explained: very few
observations have been made on these continental shelves as a whole; in large parts,
none. In some respects this makes it easy to suggest future research directions.
In shelf regions where hardly any data whatsoever are available, any new
measurements are welcome. In other respects it makes it very difficult. To identify the
key elements that need to be addressed as a matter of urgency particularly for
contiguous Third World countries with severely limited funds and research capability
is near to impossible. With these caveats, let us proceed by examining the research
needs for the coastal oceans in the sequence in which they were discussed in the
preceding sections, starting with the shelf regions of the Mozambique Channel.
The wide shelf on the eastern seaboard of Mozambique plays a recognised
important role in local fisheries and thus in the economy of the adjacent country (e.g.,
42
Sćtre and de Paula e Silva, 1979; Brinca et al., 1981). Its waters are likely to be
predominantly influenced by the reigning winds and in particular the occasional
incidence of severe cyclones. The effects of such winds on the hydrographic
characteristics, especially vertical stratification, the primary productivity and the
distribution of biota need urgent research attention. It has now been shown
conclusively (De Ruijter et al., 2002) that there is no continuous, intense Mozambique
Current that borders this shelf region, but that instead a series of eddies are formed in
the narrows of the channel that subsequently progress poleward. The influence of
these drifting eddies on the adjacent shelf is not known yet. Quartly and Srokosz
(2004) have shown that they extract water, rich in phytoplankton, from the surface
layers of the shelf. These potentially important processes need to be studied with in
situ measurements. These recent research developments also point to the need for
more, extensive deep-sea investigations.
The lack of hydrographic stations to describe and demarcate the water masses and
the circulation in the deeper parts of the Mozambique Channel (Lutjeharms, 1977)
also has an effect on what is known on the influence of the deep-sea on shelf waters.
Although recent work (e.g., De Ruijter et al., 2002; Ridderinkhof and De Ruijter,
2003) has started to fill the most obvious gaps, much remains to be done. The
hydrography and kinematics of the eastern side of the Mozambique Channel, for
instance, remain cloaked in mystery. The influence of this particular deep-sea region
on the western shelf of Madagascar therefore also remains unknown.
For the whole South West Indian Ocean this eastern side of the Mozambique
Channel is probably the coastal ocean about which least is known. At the northern tip
of Madagascar some work was done on shelf and adjacent coastal processes during
the 1960s (e.g., Angot and Ménaché, 1963; Gerard, 1964; Angot and Gerard, 1966;
Donguy and Piton, 1969), but along the central and southern parts there is to my
knowledge no adequate information whatsoever. The most basic investigations on
water masses, current patterns, biota and their respective variabilities need to be
undertaken here. This is also true for the eastern shelf of Madagascar.
As was seen in the preceding sections, only the most basic facts are know about the
two branches of the East Madagascar Current along this shelf of Madagascar. Since
the shelf here is very narrow (Figure 20.1) the fast moving waters of this current are
more than likely to have a decisive influence on the shelf waters. However, as long as
no detailed observations are made and no careful monitoring is done, this reasonable
hypothesis has to remain in the realm of speculation. Of particular importance is a
better understanding of the upwelling cell at the south-eastern corner of Madagascar.
Only one set of dedicated observations have been made here to date (Machu et al.,
2002) and much more needs to be learnt about the possible influence of this upwelling
on the biological productivity of the region. Anecdotal information from local
subsistence fishermen suggests (M. Rouault, personal communication) that it may be
a rich region for fisheries. Its investigation therefore also may hold considerable
economic and fisheries management consequences. In comparison to these shelf
regions off Madagascar and Mozambique, comparatively much more is known about
the shelf regions off South Africa.
For the shelf region inshore of the northern Agulhas Current a number of things
stand out that need further investigation if we are to understand the physico-chemical
and biological behaviour of these particular shelf waters. It has been hypothesised that
the upwelling cell at the northern tip of the Natal Bight, at St Lucia, may control the
nutrient supply to the whole bight and thus the ecosystem of this shelf province. The
43
intensity of this upwelling cell and the progress of water from there onto the shelf of
the rest of the bight need to be monitored carefully if this hypothesis is to be verified.
The shelf downstream from the bight is much narrower (Figure 20.1) and the water
movement on it therefore conceivably much simpler. The one exception to this simple
movement in sympathy with that of the Agulhas Current may come about due to the
passage of Natal Pulses.
This irregularly occurring meander on the trajectory of the Agulhas Current has
been shown to have a dramatic influence on the currents over the adjacent shelf
(Gründlingh, 1979; Lutjeharms and Connell, 1989) during its passage. To date this
has been observed only fortuitously, although it is evident in some historical current
meter records (e.g., Schumann, 1982). This shelf is the known conduit for the
migration of whales and the annual sardine run. It is unknown what effect the passage
of a Natal Pulse has on these migrations or if the unusual, but potentially beneficial,
currents associated with these current meanders are in fact purposefully used by these
animals. It would seem clear that this needs investigation. For the shelf off the
southern Agulhas Current the research questions are different.
It has been surmised (Lutjeharms and Meyer, 2004) that the upwelling cell at the
eastern end of the Agulhas Bank, adjacent to Port Alfred, may dominate the flux of
bottom water onto this shelf region. This proposition has considerable implications
for a proper understanding of the vertical stratification of the waters over the shelf,
the nutrient supply to the shelf and the ecology of this large coastal region.
Observations on this upwelling cell to date have all been inadvertent. The need for a
dedicated observational programme seems essential, particularly since the Agulhas
Bank is the spawning region for anchovy that sustain the prime pelagic fisheries of
South Africa. Such a programme should at a minimum study the nature and driving
forces of the Port Alfred upwelling cell, the movement of upwelled water from here
and the variability of the system of bottom water supply. It is not only the bottom
water that is of importance; the whole physico-chemical nature of the waters over the
Agulhas Bank is - bearing in mind its probable economic importance - amazingly
poorly known.
To date only two quasi-synoptic research cruises have been undertaken to establish
the hydrographic structure of the Agulhas Bank as a whole (Lutjeharms et al., 1981b;
1983). During these cruises only the thermal structure of the water masses was
measured. Notwithstanding the importance of this shelf region for the local fisheries,
no dedicated hydrographic cruise or set of cruises has covered the full extent of the
bank since the aforementioned cruises, nor are there immediate plans to rectify this
situation. This seems a particularly serious gap in the knowledge of these coastal
oceans that needs urgent attention.
The waters on the Agulhas Bank are influenced not only by normal solar and air-
sea interaction processes, nor only by the imbedded upwelling cell, but also by
interaction with the juxtapositioned Agulhas Current. As is evident from the above
review, plumes of warm surface water from the Agulhas Current move onto the bank
and may spread over extensive parts of it. It has consequently been surmised (Swart
and Largier, 1987) that this input of advected surface water may play an important, if
not decisive, role in maintaining the very strong vertical temperature gradients over
the bank. Attempts at quantifying this process and comparing it with the role of
insolation would be valuable and contribute to a better understanding of the factors
playing a role in the hydrography of this important part of the South African shelf and
its role in the ecology of the region, particularly the spawning of anchovy.
44
In short, even an entirely subjective listing of research priorities for the coastal
seas of the South West Indian Ocean demonstrates unequivocally the urgent need for
research in order to understand even the most fundamental, descriptive aspects of
many of the shelf regions here.
Acknowledgements
This review was undertaken during a personally difficult time. I therefore thank the
editors for their considerable patience and understanding. I wish to thank in particular
Dr Mike Roberts for kindly making data from the Mozambique Channel available to
me before publication. The volume on coastal ocean studies off Natal (Schumann,
1988a) is a veritable gold-mine of useful information for a shelf region where
investigations have virtually stopped since the commercialisation of the South African
CSIR (Lutjeharms and Thompson, 1993). Prof. Geoff Brundrit and Dr John Rogers
are thanked for a critical reading of the manuscript and for valuable comments that
made the final product better. Financial support came from an award by the National
Research Foundation of South Africa and the IDYLE programme of the French
Institut de Recherche pour le Développement.
Bibliography
Anderson, F. P., M. L. Gründlingh and C. C. Stavropoulos, 1988. Kinematics of the southern Natal
coastal circulation: some historic measurements 1962-63. S. Afr. J. Sci., 84, 857-860.
Angot, M. and R. Gerard, 1966. Charactčres hydrologiques de l'eau de surface au Centre ORSTOM de
Nosy-Bé de 1962 ŕ 1965). Cah. ORSTOM Océanogr., 4, 37-54.
Angot, M. and M. Ménaché, 1963. Premiéres données hydrologiques sur la région voisine de Nosy Bé
(nord-ouest de Madagascar). Cah. ORSTOM Océanogr., 3, 7-15.
Anonymous, 1983. Cruise report R/V "Dr. Fridtjof Nansen", Fisheries Resources Survey, Madagascar,
16-28 June 1983. Reports on Surveys with the R/V Dr Fridtjof Nansen, Institute of Marine
Research, Bergen, 9 pp.
Arhan, M., H. Mercier and J. R. E. Lutjeharms, 1999. The disparate evolution of three Agulhas rings in
the South Atlantic Ocean. J. Geophys. Res., 104, 20,987-21,005.
Asplin, L., M. D. Skogen, W. P. Budgell, V. Dove, E. Andre, T. Gammelsrřd, A. M. Hoguane, 2004.
Numerical modelling of currents and hydrography of the Mozambique Channel. In preparation.
Atkins, G. R., 1970. Wind and current patterns in False Bay. Trans. Roy. Soc. S. Afr., 39, 139- 148.
Augustyn, C. J., M. R. Lipiski, W. H. H. Sauer, M. J. Roberts and B. A. Mitchell-Innes, 1994.
Chokka squid on the Agulhas Bank: life history and ecology. S. Afr. J. Sci., 90, 143-154.
Barange, M., 1994. Acoustic identification, classification and structure of biological patchiness on the
edge of the Agulhas Bank and its relation to frontal features. S. Afr. J. Mar. Sci., 14, 333-347.
Beckley, L. E., 1983. Sea-surface temperature variability around Cape Recife, South Africa. S. Afr. J.
Sci., 79, 436-438.
Beckley, L. E., 1988. Spatial and temporal variability in sea temperature in Algoa Bay, South Africa.
S. Afr. J. Sci., 84, 67-69.
Beckley, L. E. and R. C. van Ballegooyen, 1992. Oceanographic conditions during three
ichthyoplankton surveys of the Agulhas Current in 1990/91. S. Afr. J. Mar. Sci., 12, 83-93.
Beckley, L. E. and J. D. Hewitson, 1994. Distribution and abundance of clupeoid larvae along the east
coast of South Africa in 1990/91. S. Afr. J. Mar. Sci., 14, 205-212.
Biastoch, A. and W. Krauß, 1999. The role of mesoscale eddies in the source regions of the Agulhas
Current. J. Phys. Oceanogr., 29, 2303-2317.
Biastoch, A., C. J. C. Reason, J. R. E. Lutjeharms and O. Boebel, 1999. The importance of flow in the
Mozambique Channel to seasonality in the greater Agulhas Current system. Geophys. Res. Lett.,
26, 3321-3324.
45
Boebel, O., J. Lutjeharms, C. Schmid, W. Zenk, T. Rossby and C. Barron, 2003. The Cape Cauldron: a
regime of turbulent inter-ocean exchange. Deep-Sea Res. II, 50, 57-86.
Boyd, A. J., B. B. S. Tromp and D. A. Horstman, 1985. The hydrology off the South African south-
western coast between Cape Point and Danger Point in 1975. S. Afr. J. Mar. Sci., 3, 145-168.
Boyd, A. J., J. Tauton-Clark and G. P. J. Oberholster, 1992. Spatial features of the near-surface and
midwater circulation patterns off western and southern South Africa and their role in the life
histories of various commercially fished species. S. Afr. J. Mar. Sci., 12, 189-206.
Boyd, A. J., and F. A. Shillington, 1994. Physical forcing and circulation patterns on the Agulhas
Bank. S. Afr. J. Sci., 90, 114-122.
Brinca, L., A. Jorge da Silva, L. Sousa, I. M. Sousa and R. Sćtre, 1983. A survey of the fish resources
at Sofala Bank, Mozambique. Reports on Surveys with the R/V Dr Fridtjof Nansen, Serviço de
Investigaçőes Pesqueiras, Maputo, Institute of Marine Research, Bergen, 70 + 15 pp.
Brinca, L., F. Rey, C. Silva and R. Sćtre, 1981. A survey on the marine fish resources of Mozambique.
Reports on Surveys with the R/V Dr Fridtjof Nansen, Instituto de Desenvolvimento Pesqueiro,
Maputo, Institute of Marine Research, Bergen, 58 pp.
Burchall, J., 1968. An evaluation of primary productivity studies in the continental shelf region of the
Agulhas Current near Durban (1961-1966). Investigatl Rep., Oceanogr. Res. Inst., Durban, 21, 44
pp.
Byrne, D. A., A. L. Gordon and W. F. Haxby, 1995. Agulhas eddies: a synoptic view using Geosat
ERM data. J. Phys. Oceanogr., 25, 902-917.
Carter, R. A., 1977. The distribution of calando copepods in the Agulhas Current system off Natal,
South Africa. Unpublished MSc thesis, University of Natal, 165 pp.
Carter, R. A., H. F. McMurray and J. L. Largier, 1987. Thermocline characteristics and phytoplankton
dynamics in Agulhas Bank waters. S. Afr. J. Mar. Sci., 5, 327-336.
Carter, R. and J. d'Aubrey, 1988. Inorganic nutrients in Natal continental shelf waters. In Coastal
Ocean Studies off Natal, South Africa, E. H. Schumann, ed. Lecture Notes on Coastal and Estuarine
Studies 26, Springer-Verlag, Berlin, pp. 131-151.
Carter, R. and M. H. Schleyer, 1988. Plankton distribution in Natal coastal waters. In Coastal Ocean
Studies off Natal, South Africa, E. H. Schumann, ed. Lecture Notes on Coastal and Estuarine
Studies 26, Springer-Verlag, Berlin, pp.152-177.
Chapman, P. and J. L. Largier, 1989. On the origin of Agulhas Bank bottom water. S. Afr. J. Sci., 85,
515-519.
Cooke, A., J. R. E. Lutjeharms and P. Vasseur, 2004. Marine and coastal natural history and
conservation in Madagascar. In The Natural History of Madagascar, S. M. Goodman and J. P.
Benstead, eds, The University of Chicago Press, Chicago, pp. 179-209.
Cram, D. L., 1970. A suggested origin for the cold surface water in central False Bay. Trans. Roy. Soc.
S. Afr., 39, 129-137.
Crimp, S. J., J. R. E. Lutjeharms and S. J. Mason, 1998. Sensitivity of a tropical-temperate trough to
sea-surface temperature anomalies in the Agulhas retroflection region. Water S. A., 24, 93-101.
De Decker, A. H. B., 1973. Agulhas Bank plankton. In The Biology of the Indian Ocean, B. Zeitzschel,
ed., Springer-Verlag, Berlin, pp. 189-219.
De Ruijter, W. P. M., P. J. van Leeuwen and J. R. E. Lutjeharms, 1999a. Generation and evolution of
Natal Pulses: Solitary meanders in the Agulhas Current. J. Phys. Oceanogr., 29, 3043-3055.
De Ruijter, W. P. M., A. Biastoch, S. S. Drijfhout, J. R. E. Lutjeharms, R. P. Matano, T. Pichevin, P. J.
van Leeuwen and W. Weijer, 1999b. Indian-Atlantic inter-ocean exchange: dynamics, estimation
and impact. J. Geophys. Res., 104, 20,885-20,911.
De Ruijter, W. P. M., H. Ridderinkhof, J. R. E. Lutjeharms, M. W. Schouten and C. Veth, 2002.
Observations of the flow in the Mozambique Channel. Geophys. Res. Lett., 29,
10.1029/2001GL013714.
De Ruijter, W. P. M., H. M. van Aken, E. Beier, J. R. E. Lutjeharms, R. P. Matano and M. W.
Schouten, 2004. Eddies and dipoles around South Madagascar: formation, pathways and large-
scale impact. Deep-Sea Res. I, 51, 383-400.
De Villiers, S., 1998. Seasonal and interannual variability in phytoplankton biomass on the southern
African continental shelf: evidence from satellite-derived pigment concentrations. S. Afr. J. Mar.
Sci., 19, 169-179.
DiMarco, S. F., P. Chapman and W. D. Nowlin, 2000. Satellite observations of upwelling on the
continental shelf south of Madagascar. Geophys. Res. Lett., 27, 3965-3968.
Dingle, R. V., G. V. Birch, J. M. Bremner, R. H. de Decker, A. du Plessis, J. A. Engelbrecht, M. J.
Fincham, T. Fitton, B. W. Flemming, R. I. Gentle, S. H. Goodlad, A. K. Martin, E. G. Mills, G. J.
46
Moir, R. J. Parker, S. H. Robson, J. Rogers, D. A. Salmon, W. G. Sieser, E. S. W. Simpson, C. P.
Summerhayes, F. Westall, A. Winter and M. W. Woodborne, 1987a. Bathymetry around Southern
Africa (SE Atlantic & SW Indian Oceans). Ann. S. Afr. Mus., 98, 127 (separate map1).
Dingle, R. V., G. V. Birch, J. M. Bremner, R. H. de Decker, A. du Plessis, J. A. Engelbrecht, M. J.
Fincham, T. Fitton, B. W. Flemming, R. I. Gentle, S. H. Goodlad, A. K. Martin, E. G. Mills, G. J.
Moir, R. J. Parker, S. H. Robson, J. Rogers, D. A. Salmon, W. G. Sieser, E. S. W. Simpson, C. P.
Summerhayes, F. Westall, A. Winter and M. W. Woodborne, 1987b. Deep-sea sedimentary
environments around southern Africa (South-East Atlantic and South-West Indian Oceans). Ann.
S. Afr. Mus., 98, 127 (separate map 2).
Donguy, J.-R. and B. Piton, 1969. A perçu des conditions hydrologiques de la partie nord du canal de
Mozambique. Cah. ORSTOM Océanogr., 7, 3-26.
Donguy, J.-R. and B. Piton, 1991. The Mozambique Channel revisited. Oceanologica Acta, 14, 549-
558.
Duncombe Rae, C. M., 1991. Agulhas retroflection rings in the South Atlantic Ocean; an overview. S.
Afr. J. Mar. Sci., 11, 327-344.
Duncombe Rae, C. M., F. A. Shillington, J. J. Agenbag, J. Taunton-Clark and M. L. Gründlingh, 1992.
An Agulhas Ring in the South East Atlantic Ocean and its interaction with the Benguela upwelling
frontal system. Deep-Sea Res., 39, 2009-2027.
Duncombe Rae, C. M., S. L. Garzoli and A. L. Gordon, 1996. The eddy field of the southeast Atlantic
Ocean: a statistical census from the Benguela Sources and Transports Project. J. Geophys. Res.,
101, 11,949-11,964.
Eagle, G. A. and M. J. Orren, 1985. A seasonal investigation of the nutrients and dissolved oxygen in
the water column along two lines of stations south and west of South Africa. Nat. Res. Inst.
Oceanol., CSIR. Res. Rep., 567: 52 pp.
Fine, R. A., M. J. Warner and R. F. Weiss, 1988. Water mass modification of the Agulhas
retroflection: chlorofluoromethane studies. Deep-Sea Res., 35, 311-332.
Flemming, B. W., 1978. Underwater sand dunes along the southeast African continental margin -
observations and implications. Mar. Geol., 26, 177-198.
Flemming, B. W., 1980. Sand transport and bedform patterns on the continental shelf between Durban
and Port Elizabeth (Southeast African Continental Margin). Sediment. Geol., 26, 179-205.
Flemming, B. W., 1981. Factors controlling shelf sediment dispersal along the southeast African
continental margin. Mar. Geol. 42, 259-277.
Flemming, B. and R. Hay, 1988. Sediment distribution and dynamics of the Natal continental shelf. In
Coastal Ocean Studies off Natal, South Africa, E. H. Schumann, ed., Lecture Notes on Coastal and
Estuarine Studies 26, Springer-Verlag, Berlin, pp. 47-80.
Fowler, J. L. and A. J. Boyd, 1998. Transport of anchovy and sardine eggs and larvae from the western
Agulhas Bank to the west coast during the 1993/94 and 1994/95 spawning seasons. S. Afr. J. Mar.
Sci., 19, 181-195.
Garratt, P. A., 1988. Notes on seasonal abundance and spawning of some important offshore linefish in
Natal and Transkei waters, southern Africa. S. Afr. J. Mar. Sci., 7, 1-8.
Garzoli, S. L., A. L. Gordon, V. Kamenkovich, D. Pillsbury and C. Duncombe-Rae (sic), 1996.
Variability and sources of the southeastern Atlantic circulation. J. Mar. Res., 54, 1039-1071.
Gerard, R., 1964. Étude de l'eau de mer de surface dans une baie de Nosy-Bé. Cah. ORSTOM
Océanogr., 2, 5-26.
Gill, A. E., 1977. Coastally trapped waves in the atmosphere. Quart.J. Roy.Meteorol. Soc., 103, 431-
440.
Gill, A. E. and E. H. Schumann, 1979. Topographically induced changes in the structure of an inertial
coastal jet: application to the Agulhas Current. J. Phys. Oceanogr., 9, 975-991.
Gordon, A. L., J. R. E. Lutjeharms and M. L. Gründlingh, 1987. Stratification and circulation at the
Agulhas Retroflection. Deep-Sea Res., 34, 565-599.
Goschen, W. S. and E. H. Schumann, 1988. Ocean current and temperature structures in Algoa Bay
and beyond in November 1986. S. Afr. J. Mar. Sci., 7, 101-116.
Goschen, W. S. and E. H. Schumann, 1990. Agulhas Current variability and inshore structures off the
Cape Province, South Africa. J. Geophys. Res., 95, 667-678.
Goschen, W. S. and E. H. Schumann, 1994. An Agulhas Current intrusion into Algoa Bay during
August 1988. S. Afr. J. Mar. Sci., 14, 47-57.
Goschen, W. S. and E. H. Schumann, 1995. Upwelling and the occurrence of cold water around Cape
Recife, Algoa Bay, South Africa. S. Afr. J. Mar. Sci., 16, 57-67.
47
Gründlingh, M. L., 1974. A description of inshore current reversals off Richard's Bay based on
airborne radiation thermometry. Deep-Sea Res., 21, 47-55.
Gründlingh, M. L., 1979. Observation of a large meander in the Agulhas Current. J. Geophys. Res.,
84, 3776-3778.
Gründlingh, M. L., 1983. On the course of the Agulhas Current. S. Afr. Geograph. J., 65, 49-57.
Gründlingh, M. L. and A. F. Pearce, 1984. Large vortices in the northern Agulhas Current. Deep-Sea
Res., 31, 1149-1156.
Gründlingh, M. L., 1986. Features of the Northern Agulhas Current in spring, 1983. S. Afr. J. Sci., 82,
18-20.
Gründlingh, M. L., I. T. Hunter and E. Potgieter, 1989. Bottom currents at the entrance to False Bay,
South Africa. Cont. Shelf Res., 9, 1029-1048.
Gründlingh, M. L. and A. F. Pearce, 1990. Frontal features of the Agulhas Current in the Natal Bight.
S. Afr. Geograph. J., 72, 11-14.
Gründlingh, M. L. and J. L. Largier, 1991. Physical oceanography of False Bay: a review. Trans. Roy.
Soc. S. Afr., 47, 387-400.
Gründlingh, M. L., 1992a. Agulhas Current meanders: review and a case study. S. Afr. Geogr. J., 74,
19-28.
Gründlingh, M. L., 1992b. Quasi-synoptic survey of the thermohaline properties of False Bay. S. Afr.
J. Sci., 88, 325-334.
Harris, T. F. W., 1964. Notes on Natal coastal waters. S. Afr. J. Sci., 60, 237-241.
Harris, T. F. W., 1978. Review of coastal currents in Southern African waters. South African National
Science Programmes Report, CSIR Rep. 30, vii + 103 pp.
Heydorn, A. E. F., N. D. Bang, A. F. Pearce, B. W. Flemming, R. A. Carter, M. H. Schleyer, P. F.
Berry, G. R. Hughes, A. J. Bass, J. H. Wallace, R. P. van der Elst, R. J. M. Crawford and P. A.
Shelton, 1978. Ecology of the Agulhas Current region: an assessment of biological responses to
environmental parameters in the South-West Indian Ocean. Trans. Roy. Soc. S. Afr., 43, 151-190.
Hughes, G. R., P. Luschi, R. Mencacci and F. Papi, 1998. The 7000-km oceanic journey of a
leatherback turtle tracked by satellite. J. Exp. Mar. Biol. Ecol., 229, 209-217.
Hunter, I. T., 1981. On the land breeze circulation of the Natal coast. S. Afr. J. Sci., 77, 376-378.
Hunter, I. T., 1987. The weather of the Agulhas Bank and Cape south coast. CSIR Res. Rep., 634, 184
pp.
Hunter, I. T., 1988. Climate and weather off Natal. In Coastal Ocean Studies off Natal, South Africa,
E. H. Schumann, ed., Lecture Notes on Coastal and Estuarine Studies 26, Springer-Verlag, Berlin,
pp. 81- 100.
Hutchings, L., 1994. The Agulhas Bank: a synthesis of available information and a brief comparison
with other east-coast shelf regions. S. Afr. J. Sci., 90, 179-185.
Japp, D. W., P. Sims and M. J. Smale, 1994. A review of the fish resources of the Agulhas Bank. S.
Afr. J. Sci., 90, 123-134.
Jorge da Silva, A., A. Mubango and R. Sćtre, 1981. Information on oceanographic cruises in the
Mozambique Channel. Revista de Investigaçăo Pesquira, 2, Instituto de Desenvolvimento
Pesqueiro, Maputo, República Popular de Moçambique, 89 pp.
Jorge da Silva, A., 1984a. Circulation system and areas of potentially successful tuna fishing with
surface methods off Mozambique. Revista de Investigaçăo Pesquira, 11, Instituto de Investigaçăo
Pesqueiro, Moçambique, pp. 5-40.
Jorge da Silva, A., 1984b. Report on the oceanographic investigations carried out at the Sofala Bank
by the Soviet trawler "Sevastopolsky Rybak" in September-December 1982. Revista de
Investigaçăo Pesquira, 10, Instituto de Investigaçăo Pesqueiro, Maputo, Moçambique, pp. 5-35.
Jorge da Silva, A., 1984c. Hydrology and fish distribution at the Sofala Bank (Mozambique). Revista
de Investigaçăo Pesquira, 12, Instituto de Investigaçăo Pesqueiro, Moçambique, pp. 5-36.
Jury, M. R., 1988. A climatological mechanism for wind-driven upwelling near Walker Bay and
Danger Point, South Africa. S. Afr. J. Mar. Res., 6, 175-181.
Jury, M. (R.) and N. Walker, 1988. Marine boundary layer modification across the edge of the Agulhas
Current. J. Geophys. Res., 93, 647-654.
Jury, M. (R.), 1991. The weather of False Bay. Trans. Roy. Soc. S. Afr., 47, 401-417.
Jury, M. R. and B. Pathack, 1991. A study of climate and weather variability over the tropical
southwestern Indian Ocean. Meteor. Atmos. Phys., 47, 37-48.
Jury, M. R., H. R. Valentine and J. R.E. Lutjeharms, 1993. Influence of the Agulhas Current on
summer rainfall on the southeast coast of South Africa. J. Appl. Meteorol., 32, 1282-1287.
48
Jury, M. R., 1994. A review of the meteorology of the eastern Agulhas Bank. S. Afr. J. Sci., 90, 109-
113.
Largier, J. L. and V. P. Swart, 1987. East-west variation in thermocline breakdown on the Agulhas
Bank. S. Afr. J. Mar. Sci., 5, 263-272.
Largier, J. L., P. Chapman, W. T. Peterson and V. P. Swart, 1992. The western Agulhas Bank:
circulation, stratification and ecology. S. Afr. J. Mar. Sci., 12, 319-339.
Lee-Thorp, A. M., M. Rouault and J. R. E. Lutjeharms, 1998. Cumulus cloud formation above the
Agulhas Current. S. Afr. J. Sci., 94, 351-354.
Lee-Thorp, A. M., M. Rouault and J. R. E. Lutjeharms, 1999. Moisture uptake in the boundary layer
above the Agulhas Current: a case study. J. Geophys. Res., 104, 1423-1430.
Luschi, P., A. Sale, R. Mencacci, G. R. Hughes, J. R. E. Lutjeharms and F. Papi, 2003. Current
transport of leatherback sea turtles (Dermochelys coriacea) in the ocean. Proc. Royal Soc. London,
Ser. B Biol. Scis, 270 (Supplement): S129-S132.
Lutjeharms, J. R. E., 1977. The need for oceanologic research in the South-West Indian Ocean. S. Afr.
J. Sci., 73, 40-43.
Lutjeharms, J. R. E., N. D. Bang and C. P. Duncan, 1981a. Characteristics of the currents east and
south of Madagascar. Deep-Sea Res., 28, 879-899.
Lutjeharms, J. R. E., N. D. Bang and H. R. Valentine, 1981b. Die fisiese oseanologie van die
Agulhasbank. Deel I: Vaart 170 van die N.S. Thomas B. Davie. WNNR Navorsingsverslag 386, 38
pp.
Lutjeharms, J. R. E. and H. R. Valentine, 1983. Die fisiese oseanologie van die Agulhasbank. Deel 2:
Vaart 185 van die N.S. Thomas B. Davie. WNNR Navorsingsverslag 557, 15 pp.
Lutjeharms, J. R. E. and N. M. Walters, 1985. Ocean colour and thermal fronts south of Africa. In
South African Ocean Colour and Upwelling Experiment, L. V. Shannon, ed., Sea Fisheries
Research Institute, Cape Town, pp. 227-237.
Lutjeharms, J. R. E., D. Baird and I. T. Hunter, 1986a. Seeoppervlak dryfgedrag aan die Suid-
Afrikaanse suidkus in 1979. S. Afr. J. Sci., 82, 324-326.
Lutjeharms, J. R. E., R. D. Mey and I. T. Hunter, 1986b. Cloud lines over the Agulhas Current. S. Afr.
J. Sci., 82, 635-640.
Lutjeharms, J. R. E. and A. L. Gordon, 1987. Shedding of an Agulhas Ring observed at sea. Nature,
325, 138-140.
Lutjeharms, J. R. E. and J. M. Meeuwis, 1987. The extent and variability of South-East Atlantic
upwelling. S. Afr. J. Mar. Sci., 5, 51-62.
Lutjeharms, J. R. E., 1988. Remote sensing corroboration of retroflection of the East Madagascar
Current. Deep-Sea Res., 35, 2045-2050.
Lutjeharms, J. R. E. and H. R. Roberts, 1988. The Natal Pulse; an extreme transient on the Agulhas
Current. J. Geophys. Res., 93, 631-645.
Lutjeharms, J. R. E. and H. R. Valentine, 1988. Evidence for persistent Agulhas rings southwest of
Cape Town. S. Afr. J. Sci., 84, 781-783.
Lutjeharms, J. R. E. and R. C. van Ballegooyen, 1988. The retroflection of the Agulhas Current. J.
Phys. Oceanogr., 18, 1570-1583.
Lutjeharms, J. R. E. and A. Jorge da Silva, 1988. The Delagoa Bight eddy. Deep-Sea Res., 35, 619-
634.
Lutjeharms, J. R. E. and A. D. Connell, 1989. The Natal Pulse and inshore counter currents off the
South African east coast. S. Afr. J. Sci., 85, 533-535.
Lutjeharms, J. R. E., R. Catzel and H. R. Valentine, 1989a. Eddies and other border phenomena of the
Agulhas Current. Cont. Shelf Res., 9, 597-616.
Lutjeharms, J. R. E., M. L. Gründlingh and R. A. Carter, 1989b. Topographically induced upwelling in
the Natal Bight. S. Afr. J. Sci., 85, 310-316.
Lutjeharms, J. R. E., 1991. The temperature/salinity relationships of the South West Indian Ocean. S.
Afr. Geographer, 18, 15-31.
Lutjeharms, J. R. E. and P. L. Stockton, 1991. Aspects of the upwelling regime between Cape Point
and Cape Agulhas. S. Afr. J. Mar. Sci., 10, 91-102.
Lutjeharms, J. R. E., J. Olivier and E. Lourens, 1991. Surface fronts of False Bay and vicinity. Trans.
Roy. Soc. S. Afr., 47, 433-445.
Lutjeharms, J. R. E. and J. A. Thomson, 1993. Commercializing the CSIR and the death of science.
S. Afr. J. Sci., 89, 8-14.
Lutjeharms, J. R. E., C.-T. Liu, W.-S. Chuan and C.-Z. Shyu, 1993. On some similarities between the
oceanic circulations off Southern Africa and off Taiwan. S. Afr. J. Sci., 89, 367-371.
49
Lutjeharms, J. R. E., 1996. The exchange of water between the South Indian and the South Atlantic. In
The South Atlantic: Present and Past Circulation, G. Wefer, W. H. Berger, G. Siedler and D.
Webb, eds., Springer-Verlag, Berlin, pp. 125-162.
Lutjeharms, J. R. E. and J. Cooper, 1996. Interbasin leakage through Agulhas Current filaments. Deep-
Sea Res. I, 43, 213-238.
Lutjeharms, J. R. E. and W. P. M. de Ruijter, 1996. The influence of the Agulhas Current on the
adjacent coastal zone: possible impacts of climate change. J. Mar. Syst., 7, 321-336.
Lutjeharms, J. R. E., A. A. Meyer, I. J. Ansorge, G. A. Eagle and M. J. Orren, 1996. The nutrient
characteristics of the Agulhas Bank. S. Afr. J. Mar. Sci., 17, 253-274.
Lutjeharms, J. R. E., 1998. Coastal hydrography. In A Field Guide to the Eastern and Southern Cape
Coast, R. Lubke and I. de Moor, eds, The Wildlife and Environment Society of Southern Africa,
Grahamstown, University of Cape Town Press, Rondebosch, pp. 50-61.
Lutjeharms, J. R. E. and E. Machu, 2000. An upwelling cell inshore of the East Madagascar Current.
Deep-Sea Res. I, 47, 2405-2411.
Lutjeharms, J. R. E., J. Cooper and M. Roberts, 2000a. Upwelling at the inshore edge of the Agulhas
Current. Cont. Shelf Res., 20, 737-761.
Lutjeharms, J. R. E., P. M. Wedepohl and J. M. Meeuwis, 2000b. On the surface drift of the East
Madagascar and the Mozambique Currents. S. Afr. J. Sci., 96, 141-147.
Lutjeharms, J. R. E., H. R. Valentine and R. C. van Ballegooyen, 2000c. The hydrography and water
masses of the Natal Bight, South Africa. Cont. Shelf Res., 20, 1907-1939.
Lutjeharms, J. R. E., W. P. M. de Ruijter, H. Ridderinkhof, H. van Aken, C. Veth, P. J. van Leeuwen,
S. S. Drijfhout, J. H. F. Jansen and G.-J. A. Brummer, 2000d. MARE and ACSEX: new research
programmes on the Agulhas Current system. S. Afr. J. Sci., 96, 105-110.
Lutjeharms, J. R. E. and I. Ansorge, 2001. The Agulhas Return Current. J. Mar. Syst., 30, 115-138.
Lutjeharms, J. R. E., P. Penven and C. Roy, 2003a. Modelling the shear edge eddies of the southern
Agulhas Current. Cont. Shelf Res., 23, 1099-1115.
Lutjeharms, J. R. E., O. Boebel and H. T. Rossby, 2003b. Agulhas cyclones. Deep-Sea Research II,
50(1): 35-56.
Lutjeharms, J. R. E. and A. A. Meyer, 2004. The origin and circulation of bottom water on the Agulhas
Bank, South Africa. Cont. Shelf Res., in preparation.
Machu, E., J. R. E. Lutjeharms, A. M. Webb and H. M. van Aken, 2002. First hydrographic evidence
of the south-east Madagascar upwelling cell. Geophys. Res. Lett., 29, doi:10.1029/2002GL015381.
Malan, O. G. and E. H. Schumann,1979. Natal shelf circulation revealed by Landsat imagery. S.Afr. J.
Sci., 75, 136-137.
Maltrud, M. E., R. D. Smith, A. J. Semtner and R. C. Malone, 1998. Global eddy-resolving ocean
simulations driven by1985-1995 atmospheric winds. J. Geophys. Res., 103, 30,825-30,853.
Martin, A. K., 1981.The influence of the Agulhas Current on the physiographic development of the
northernmost Natal Valley (SW Indian Ocean). Mar. Geol, 39, 259-276.
Martin, A. K., and B. W. Flemming, 1988. Physiography, structure and geological evolution of the
Natal continental shelf. In Coastal Ocean Studies off Natal, South Africa, E. H. Schumann, ed.,
Lecture Notes on Coastal and Estuarine Studies 26, Springer-Verlag, Berlin, pp. 11-46.
Martin, J., P. Guibout, M. Crepon and J.-C. Lizaray, 1965. Circulation superficielle dans l'ocean
Indien. Résultats de mesures faites ŕ électrodes remorguées G.E.K. entre 1955-1963. Cah.
Oceanogr., 17, suppl. 3, 221-241.
McClurg, T. M., 1988. Benthos of the Natal continental shelf. In Coastal Ocean Studies off Natal,
South Africa, E. H. Schumann, ed. Lecture Notes on Coastal and Estuarine Studies 26, Springer-
Verlag, Berlin, pp. 178-208.
McMurray, H. F., R. A. Carter and M. I. Lucas, 1993. Size-fractionated phytoplankton production in
western Agulhas Bank continental shelf waters. Cont. Shelf Res., 13, 307-329.
Menaché, M., 1963. Premičre compagne océanographique du Commandant Robert Giraud dans le
canal de Mozambique, 11 octobre au 28 novembre 1957. Cah. Oceanogr., 15, 224-235.
Mey, R. D., N. D. Walker and M. R. Jury, 1990. Surface heat fluxes and marine boundary layer
modification in the Agulhas retroflection region. J. Geophys. Res., 95, 15,997-16,015.
Meyer, A. A., J. R. E. Lutjeharms and S. de Villiers, 2002. The nutrient characteristics of the Natal
Bight, South Africa. J. Mar. Res., 35, 11-37.
Michaelis, G., 1923. Die Wasserbewegung an der Oberfläche des Indischen Ozeans im Januar und Juli.
Veröffentl. Inst. Meeresk. Uni. Berlin, n.f. A8, 32 pp.
Mitchell-Innes, B. A., A. J. Richardson and S. J. Painting, 1999. Seasonal changes in phytoplankton
biomass on the western Agulhas bank, South Africa. S. Afr. J. Mar. Sci., 21, 217-233.
50
Nehring, D. ed., 1984. The oceanological conditions in the western part of the Mozambique Channel in
February-March 1980. Geodät. geophys. Veröffentl., 4, 163 pp.
Nehring, D., E. Hagen, A. Jorge da Silva, R. Schemainda, G. Wolf, N. Michelchen, W. Kaiser, L.
Postel, F. Gosselck, U. Brenning, E. Kühner, G. Arlt, H. Siegel, L. Gohs and G. Bublitz, 1987.
Results of oceanological studies in the Mozambique Channel in February March 1980. Beitr.
Meereskd., 56, 51-63.
Oliff, W. D., 1973. Chemistry and productivity at Richard's Bay. NPRL Oceanogr. Div. Contract
Report, CFIS 37B, Durban, South Africa.
Pearce, A. F., 1977a. Some features of the upper 500 m of the Agulhas Current. J. Mar. Res., 35, 731-
753.
Pearce, A. F., 1977b. The shelf circulation off the east coast of South Africa. Council for Scientific and
Industrial Research, CSIR Res. Rep., 361, 220 pp.
Pearce, A. F., 1978. Seasonal variations of temperature and salinity on the northern Natal continental
shelf. S. Afr. Geogr. J., 60, 135-143.
Pearce, A. F., E. H. Schumann and G. S. H. Lundie, 1978. Features of the shelf circulation off the
Natal coast. S. Afr. J. Sci., 74, 328-331.
Penven, P., J. R. E. Lutjeharms, P. Marchesiello, S. J. Weeks and C. Roy, 2001a. Generation of
cyclonic eddies by the Agulhas Current in the lee of the Agulhas Bank. Geophys. Res. Lett., 26,
1055-1058.
Penven, P., C. Roy, G. B. Brundrit, A. Colin de Verdičre, P. Fréon, A. S. Johnson, J. R. E. Lutjeharms
and F. A. Shillington, 2001b. A regional hydrodynamic model of the Southern Benguela. S. Afr. J.
Sci., 97, 472 - 475.
Piton, B., and J. F. Poulain, 1974. Résultats de mesures de courants superficiels au G.E.K. effectuées
avec N.O. Vauban dans le sud-ouest de l'océan Indien (1973-1974). Office de la Recherche
Scientifique et Technique Outre-mer, Documents Scientifique de la Mission de Nosy-Bé, 47, 14 pp.
Poolman, E. and D. Terblanche, 1984. Tropical cyclones Domoina and Imboa. S. Afr.Weather Bureau
Newsl., 420, 37-46.
Probyn, T. A., B. A. Mitchell-Innes, P. C. Brown, L. Hutchings and R. A. Carter, 1994. A review of
primary production and related processes on the Agulhas Bank. S. Afr. J. Sci., 90, 166-173.
Pugh, D. T., 1987. Tides, Surges and Mean Sea Level. John Wiley and Sons, New York, 472 pp.
Quartly, G. D. and M. A. Srokosz, 2004. Eddies in the southern Mozambique Channel. Deep-Sea Res.
II, 51, 69-83.
Reason, C. J. C., 2001. Evidence for the influence of the Agulhas Current on regional atmospheric
circulation patterns. J. Climate, 14, 2769-2778.
Ridderinkhof, H., J. R. E. Lutjeharms and W. P. M. de Ruijter, 2001. A research cruise to investigate
the Mozambique Current. S. Afr. J. Sci., 97, 461 - 464.
Ridderinkhof, H., and W. P. M. de Ruijter, 2003. Moored current observations in the Mozambique
Channel. Deep-Sea Res. II, 50, 1933-1955.
Roel, B. A., J. Hewitson, S. Kerstan and I. Hampton, 1994. The role of the Agulhas Bank in the life
cycle of pelagic fish. S. Afr. J. Sci., 90, 185-196.
Rogers, J. and J. M. Bremner, 1991. The Benguela Ecosystem. Part VII. Marine-geological aspects.
Oceanogr. Mar. Biol. Annu. Rev., 29, 1-85.
Rouault, M., A. M. Lee-Thorp, I. Ansorge and J. R. E. Lutjeharms, 1995. Agulhas Current Air-Sea
Exchange Experiment. S. Afr. J. Sci., 91, 493-496.
Rouault, M., A. M. Lee-Thorp and J. R. E. Lutjeharms, 2000. The atmospheric boundary layer above
the Agulhas Current during alongcurrent winds. J. Phys. Oceanogr., 30, 40-50.
Rouault, M., S. A. White, C. J. C. Reason and J. R. E. Lutjeharms and I. Jobard, 2002. Ocean-
atmosphere interaction in the Agulhas Current and a South African extreme weather event. Weather
Forecast., 17, 655-669.
Sćtre, R. and R. de Paula e Silva, 1979. The marine fish resources of Mozambique. Reports on surveys
with the R/V Dr Fridtjof Nansen, Serviço de Investigaçőes Pesqueiras, Maputo, Institute of Marine
Research, Bergen, 179 pp.
Sćtre, R. and A. Jorge da Silva, 1982. Water masses and circulation of the Mozambique Channel.
Revista de Investigaçăo Pesquira, 3, Instituto de Desenvolvimento Pesqueiro, Maputo, República
Popular de Moçambique, 83 pp.
Sćtre, R. and A. Jorge da Silva, 1984. The circulation of the Mozambique Channel. Deep-Sea Res., 31,
485-508.
Sćtre, R., 1985. Surface currents in the Mozambique Channel. Deep-Sea Res., 32, 1457-1467.
51
Schemainda, R. and E. Hagen, 1983. On steady state intermediate vertical currents induced by the
Mozambique Current. Océanogr. Trop., 18, 81-88.
Schott, F., M. Fieux, J. Kindle, J. Swallow and R. Zantopp, 1988. The boundary currents east and
north of Madagascar. Part II. Direct measurements and model comparisons. J. Geophys. Res., 93,
4963-4974.
Schouten, M. W., W. P. M. de Ruijter, P. J. van Leeuwen and J. R. E. Lutjeharms, 2000. Translation,
decay and splitting of Agulhas rings in the south-eastern Atlantic ocean. J. Geophys. Res., 105,
21,913-21,925.
Schouten, M. W., W. P. M. de Ruijter and P. J. van Leeuwen, 2002. Upstream control of Agulhas ring
shedding. J. Geophys. Res., doi: 10.1029/2001JC000804.
Schumann, E. H., 1981. Low frequency fluctuations off the Natal coast. J. Geophys. Res., 86, 6499-
6508.
Schumann, E. H., 1982. Inshore circulation of the Agulhas Current off Natal. J. Mar. Res., 40, 43-55.
Schumann, E. H. and L.-A. Perrins, 1982. Tidal and inertial currents around South Africa. In
Proceedings of the Eighteenth International Coastal Engineering Conference, American Society of
Civil Engineers, Cape Town, South Africa, Nov. 14-19, 1982, pp. 2562-2580.
Schumann, E. H., L.-A. Perrins and I. T. Hunter, 1982. Upwelling along the south coast of the Cape
Province, South Africa. S. Afr. J. Sci., 78, 238-242.
Schumann, E. H. and L. J. Beekman, 1984. Ocean temperature structures on the Agulhas Bank. Trans.
Roy. Soc. S. Afr., 34, 191-203.
Schumann, E. H., 1986. The bottom boundary layer inshore of the Agulhas Current off Natal in August
1975. S. Afr. J. Mar. Sci., 4, 93-102.
Schumann, E. H., 1987. The coastal ocean off the east coast of South Africa. Trans. Roy. Soc. S. Afr.,
46, 215-229.
Schumann, E. H. (editor), 1988a. Coastal Ocean Studies off Natal, South Africa, Lecture Notes on
Coastal and Estuarine Studies 26, Springer-Verlag, Berlin, 271 pp.
Schumann, E. H., 1988b. Physical oceanography off Natal. In Coastal Ocean Studies off Natal, South
Africa, E. H. Schumann, ed., Lecture Notes on Coastal and Estuarine Studies 26, Springer-Verlag,
Berlin, pp. 101-130.
Schumann, E. H. and I. Li (sic) van Heerden, 1988. Observations of Agulhas Current frontal features
south of Africa. Deep-Sea Res., 35, 1355-1362.
Schumann, E. H., G. J. B. Ross and W. S. Goschen, 1988. Cold water events in Algoa Bay and along
the Cape south coast, South Africa, in March/April 1987. S. Afr. J. Sci., 84, 579-584.
Schumann, E. H., 1989. The propagation of air pressure and wind systems along the South African
coast. S. Afr. J. Sci., 85, 382-385.
Schumann, E. H. and J. A. Martin, 1991. Climatological aspects of the coastal wind field at Cape
Town, Port Elizabeth and Durban. S. Afr. Geogr. J., 73, 48-51.
Schumann, E. H., W. K. Illenberger and W. S. Goschen, 1991. Surface winds over Algoa Bay. S. Afr.
J. Sci., 87, 202-207.
Schumann, E. H., 1992. Interannual wind variability on the south and east coasts of South Africa. J.
Geophys. Res., 97, 20,397-20,403.
Schumann, E. H., A. L. Cohen and M. R. Jury, 1995. Coastal sea surface temperature variability along
the south coast of South Africa and the relationship to regional and global climate. J. Mar. Res., 53,
231-248.
Schumann, E. H., 1998. The coastal ocean off southeast Africa, including Madagascar. In The Sea,
Volume 11, Chapter 19, A. R. Robinson and K. H. Brink, eds, John Wiley & Sons, pp. 557-581.
Schumann, E. H., 1999. Wind-driven mixed layer and coastal upwelling processes off the south coast
of South Africa. J. Mar. Res., 57, 671-691.
Shannon, L. V. and P. Chapman, 1983. Suggested mechanism for the chronic pollution by oil of
beaches east of Cape Agulhas, South Africa. S. Afr. J. Mar. Sci., 1, 231-244.
Shannon, L. V., P. Chapman, G. A. Eagle and T. P. McClurg, 1983. A comparative study of tar ball
distribution and movement in two boundary current regimes. Oil and Petrochemical Pollution, 1,
243-259.
Sidorn, J. R., D. G. Bowers and A. M. Hoguane, 2001. Detecting the Zambezi river plume using
observed optical properties. Mar. Pollut. Bull., 42, 942-950.
Simpson, E. S. W., 1974. Southeast Atlantic and Southwest Indian Oceans. Chart 125A, bathymetry.
Smale, M. J., N. T. Klages, J. H. M. David and V. G. Cockroft, 1994. Predators of the Agulhas Bank.
S. Afr. J. Sci., 90, 135-142.
52
Stramma, L. and J. R. E. Lutjeharms, 1997. The flow field of the subtropical gyre of the South Indian
Ocean. J. Geophys. Res., 102, 5513-5530.
Swallow, J. C., M. Fieux and F. Schott, 1988. The boundary currents east and north of Madagascar.
Part I. Geostrophic currents and transports. J. Geophys. Res., 93, 4951-4962.
Swart, V. P. and J. L. Largier, 1987. Thermal structure of Agulhas Bank water. S. Afr. J. Mar. Sci., 5,
243-253.
Taljaard, S., 1991. The origin and distribution of dissolved nutrients in False Bay. Trans. Roy. Soc. S.
Afr., 47, 483- 493.
Tripp, R. T., (1967). An Atlas of Coastal Surface Drifts; Cape Town to Durban. South African
Oceanographic Data Centre, Department of Oceanography, University of Cape Town, Rondebosch,
South Africa, 12 pp.
Van der Elst, R. P., 1988. Shelf ichthyofauna off Natal. In Coastal Ocean Studies off Natal, South
Africa, E. H. Schumann, ed., Lecture Notes of Coastal and Estuarine Studies, 26, Springer-Verlag,
Berlin, pp. 209-225.
Van Heerden, J. and J. J. Taljaard, 1998. Africa and surrounding waters. In Meteorology and the
Southern Hemisphere, D. J. Karoly and D. G. Vincent, eds, Meteorological Monographs, 27(49):
141-174.
Van Leeuwen, P. J., W. P. M. de Ruijter and J. R. E. Lutjeharms, 2000. Natal Pulses and the formation
of Agulhas rings. J. Geophys. Res., 105, 6425-6436.
Verheye, H. M., L. Hutchings. J. A. Huggett, R. A. Carter, W. T. Peterson and S. J. Painting, 1994.
Community structure, distribution and tropic ecology of zooplankton on the Agulhas Bank with
special reference to copepods. S. Afr. J. Sci., 90, 154-165.
Walker, N. D., 1986. Satellite observations of the Agulhas Current and episodic upwelling south of
Africa. Deep-Sea Res., 33, 1083-1106.
Walker, N. D., 1990. Links between South African summer rainfall and temperature variability of the
Agulhas and Benguela Current Systems. J. Geophys. Res., 95, 3297-3319.
53
Document Outline